
Abstract
A succession of clays with thin shell beds from the French Ardennes, dated to the late Early Pliensbachian Davoei Zone and yielding abundant well-preserved remains of all five extant classes of echinoderms, is described here as a new Lagerstätte. The echinoderms occur in the form of isolated ossicles, semi-articulated plate aggregates and abundant intact skeletons with even the most delicate appendices preserved in place and the finest skeletal structures hardly blurred by recrystallisation. The outstandingly good preservation of the echinoderm specimens allows for a morphological assessment of the represented taxa in detail only rarely achievable by fossil material. The material described herein thus significantly contributes to a better understanding of the systematic position and phylogenetic background of many Early Jurassic echinoderm taxa. The echinoderms are considered to have been buried as an autochthonous or at least parautochthonous assemblage among clumps and beds of Modiolus bivalves. The depositional setting most likely represented a relatively shallow, near-shore soft-bottom environment in which conditions repeatedly favoured the settlement and subsequent effective burial of bivalve-echinoderm assemblages, leading to the formation of the thin highly fossiliferous shell beds intercalating the otherwise nearly sterile clays. The new Lagerstätte opens a unique window into the palaeoecology of an extinct shallow-water soft-bottom community closely comparable to recent analogues.
Similar content being viewed by others

Introduction
Fossil Lagerstätten, deposits with exceptionally well-preserved and/or abundant fossil occurrences, are highly important in palaeontology as they provide insights into extinct communities which are generally not obtained from normal deposits. Extraordinary preservation of fossils, especially of soft-tissue and articulated skeletons, however, requires particular combinations of physical and chemical conditions (Seilacher et al. 1985), making Konservat-Lagerstätten a relatively rare geological phenomenon. Considering the rapid post-mortem disintegration of the multisclerite echinoderm skeleton under normal depositional conditions (Ausich 2001), it is not surprising that the occurrence of articulated echinoderms is mostly bound to Konservat-Lagerstätten.
Lower Jurassic strata of NW-Europe are among the most productive in terms of articulated echinoderms. Lagerstätten like the Toarcian Posidonia-shale (Seilacher et al. 1985), the Hettangian insect marls (Heer 1865) and the Pliensbachian Starfish Beds (Goldring and Stephenson 1972) have produced well-preserved echinoderm assemblages which added considerably to the understanding of the geological history of the group. Here, we describe a new Lagerstätte dominated by echinoderms which was recently discovered in Lower Jurassic argillite successions outcropping near Sedan in the French Ardennes. Among the array of Early Jurassic occurrences of articulated echinoderms hitherto known, the Sedan Lagerstätte stands out due to, even for Konservat-Lagerstätten, the extraordinarily good preservation of the highly abundant specimens. Even the finest skeletal structures are pristinely preserved. Furthermore, many of the echinoderm taxa encountered occur both as intact skeletons, as semi-articulated plate assemblages and in the form of isolated ossicles, in most cases all entirely extractable from the argillites, which allows for a wealth of hard-part morphological information nearly equalling the standards of echinoderm neontology.
First investigations on the Sedan locality, performed shortly after its discovery in 2002, resulted in a systematic description of part of the ophiuroid fauna and included a preliminary palaeoecological model for the depositional setting (Thuy 2010). The present report is based on new observations on a more extensive outcrop area and considerably larger amounts of material that were available at the time of the first investigations, and thus allows complementing the existing data on the geological background, the accompanying fauna and the depositional setting. It aims at a first general presentation of the Lagerstätte, its echinoderm assemblage and the perspectives for future research that arise from the extraordinary preservation and abundance of the material.
The material described and illustrated herein has been deposited in the collections of the Muséum National d’Histoire Naturelle in Paris (MNHN), the Musée national d’histoire naturelle in Luxembourg (MnhnL), the Geoscientific Museum of the University of Göttingen (GZG), and the Natural History Museum in London (NHM).
Geological background
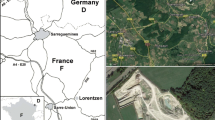
The echinoderm-bearing horizons occur within a series of dark grey clays exposed on the southern slope of the N43 road cuttings and a cutting to the north side of the road in the premises of a lorry park, south–west of Sedan in the French Ardennes (Fig. 1). The northern section (“lorry park”, Fig. 2) is stratigraphically lower, and includes nearly 4 m of dark grey clays, containing up to six irregularly developed beds of red–brown siderite nodules, each associated with thin lenses of shelly clay, containing abundant Modiolus Lamarck, 1799. A horizon nearly 2 m above the top of the section contained particularly abundant echinoderm material. The southern section is stratigraphically higher, and includes nearly 5 m of similar clays with sideritic nodule layers (Fig. 2). The lowest of these (A) was extensively sampled. The precise correspondence between the two sections is uncertain. The clays are bioturbated, and only the thin shell beds are fossiliferous.

Locality map with the position of the Sedan sections indicated by a star

Lithostratigraphy of the two studied sections with assumed correlation and position of the sampled levels. Thickness of the sections is indicated in metres
Macrofossils, in particular the echinoderm remains described herein, almost exclusively occur in the shelly beds. Bivalves of the genus Modiolus are the dominant component of the shelly beds, and are accompanied by scattered specimens of small, thin-shelled pectinid, ostreid and large inoceramid bivalves. Belemnites and ammonites are not rare, in contrast to initial observations made at a time when only a small part of the sections was accessible (Thuy 2010). The assignment to the late early Pliensbachian Davoei Zone as initially suggested by minute ammonites with affinities to the genus Androgynoceras Hyatt, 1867 (Thuy 2010) is now corroborated by new finds of typical specimens of Prodactylioceras davoei (Sowerby, 1822). Microfossils other than echinoderm remains from sieving residues of the shelly beds include benthic foraminifers, ostracods, juvenile bivalves, small cephalaspid and other gastropods, lingulid brachiopods, bryozoans, onychites (Coleoidea) and actinopterygian teeth and scales.
The material
The remains of all five extant classes of echinoderms have been abundantly retrieved from the shelly beds of the Sedan Lagerstätte, collected both by surface picking in the field of the weathered clays for the larger articulated specimens and fragments, and by screen washing of bulk sediment samples for smaller articulated specimens and isolated ossicles. Except for holothuroids, which are so far only represented by isolated sclerites and calcareous ring elements, all echinoderm groups are known from articulated specimens.
Crinoids
The most abundant group are the crinoids, leaving numerous well-preserved, articulated series of columnals and brachials on the weathered clay surface and in the sieving residues. Intact crowns with associated stalk fragments occasionally occur (Fig. 3a–c). The vast majority of the crinoid remains are attributable to the isocrinid Balanocrinus gracilis (Charlesworth, 1847) (Hess 2010). Additionally, sieving residues have produced rare dissociated remains of a yet unidentified comatulid crinoid.

Balanocrinus gracilis (Charlesworth, 1847); a proximal stem, cup and base of arms (GZG.INV.30561), lateral view; scale bar equals 1 mm. b, c cup (GZG.INV. 30562), oral (b) and aboral (c) view; scale bar equals 1 mm. dPalaeocoma milleri (Phillips, 1829), articulated specimen (GZG.INV.30553), dorsal view, as found on weathered clay surface on the lorry park cutting; scale bar equals 1 mm. eEirenura papillata Thuy, 2010, articulated disc with arm fragment (GZG.INV.30554), ventral view; scale bar equals 500 μm. fInexpectacantha acrobatica Thuy, 2010, articulated disc with arm fragments (MNHN A30315), ventral view; scale bar equals 500 μm. gSinosura brodiei (Wright, 1866), articulated specimen (GZG.INV.30555), dorsal view; scale bar equals 500 μm. All from the Pliensbachian Davoei Zone of Sedan, Ardennes, France; main echinoderm level, lorry park cutting (a–c, f) and level A, road cutting (d, e, g)
Ophiuroids
The second most abundant group, in particular in terms of articulated specimens, are the ophiuroids. A first study on part of the ophiuroid fauna revealed that as little as 4 kg of bulk sediment from a single shelly bed, collected in 2002, yielded 39 articulated discs and disc fragments and several hundreds of articulated arm fragments (Thuy 2010). Large-scale bulk sampling of the shelly beds has since raised the number of articulated discs to several hundred specimens, most of them exposing both ventral and dorsal sides. The two most common ophiuroid forms turned out to be the small-sized ophiacanthid Inexpectacantha acrobatica Thuy, 2010 (Fig. 3f) and the ophiurid Eirenura papillata Thuy, 2010 (Fig. 3e).
Other species, of which only a few specimens were available at the time of the first study, are now represented by several articulated individuals and numerous dissociated plates, allowing for a detailed systematic assessment. These include the large species Palaeocoma milleri (Phillips, 1829) (Fig. 3d), Sinosura brodiei (Wright, 1866) (Figs. 3g, 4a–b), one species comparable with Ophiomusium murravii (Forbes, 1844) (Fig. 4c–e) from the Pliensbachian of Yorkshire, UK, and Ophiura? astonensis Hess, 1964, so far only known from isolated lateral arm plates. The material is so well preserved that a systematic reassessment of these extinct taxa, closely following the standards of modern ophiuroid hard-part morphology, is possible. In addition, a single articulated specimen documents the presence in the Sedan assemblage of a yet undescribed, enigmatic species that is unlike any brittle-star form currently known from Jurassic strata (Fig. 4f). The disc of the small specimen is conspicuously dominated by a circle of 10 thick, knob-like radial shields, which is strongly reminiscent of forms attributed to the Triassic genus Aspiduriella Bolette, 1998. Further research will clarify whether the specimen represents a Jurassic relict species of Aspiduriella or rather documents a case of independent evolution of similar dorsal disc morphology.

aSinosura brodiei (Wright, 1866), detail of proximal arm segment (GZG.INV.30556), dorso-lateral view, showing spine articulation; scale bar equals 100 μm. bSinosura brodiei (Wright, 1866), articulated specimen with arm fragment (GZG.INV.30556), ventral view; scale bar equals 500 μm c, d, e“Ophiomusium” sp., articulated specimen with arm fragments (GZG.INV.30557), dorsal view (c), ventral view (d) and detail of ventral view (e); scale bars equal 1 mm. f Undetermined ophiuroid, articulated specimen (GZG.INV.30558), dorsal view; scale bar equals 500 μm. All from the Pliensbachian Davoei Zone of Sedan, Ardennes, France; main echinoderm level, lorry park cutting (a–e) and level A, road cutting (f)
The assignment of Jurassic ophiuroid taxa to the extant genus Ophiomusium Lyman, 1869 is untenable in most cases, as similarities with modern representatives of the genus are mostly superficial. The genus Mesophiomusium Kutscher & Jagt, 2000, introduced for extinct Ophiomusium-like species which, however, cannot be meaningfully reconciled with the neontological concept of the genus, needs reappraisal from the perspective of modern ophiuroid systematics, for which the material from the Sedan Lagerstätte provides a promising basis.
Scanning electron microscopy of the specimens from Sedan assignable to Sinosura brodiei reveals that some characters, including arm plate ornamentation, spine articulation structure and disc plating, are reminiscent of extant ophioleucinid brittle stars (Figs. 3g, 4a–b) (Madsen 1983; Martynov 2010). Smith et al. (1995) already tentatively placed the genus Sinosura Hess, 1964 in the Ophioleucinae, whereas Hess and Meyer (2008) transferred it to their new subfamily Aplocominae within the extinct Aplocomidae. The specimens from Sedan display some of the diagnostic characters in exceptional clarity, allowing for a reassessment of the higher taxonomic position of the genus Sinosura.
The residues of bulk sediment sampling produced a number of very small articulated ophiuroid discs, mostly assignable to Inexpectacantha acrobatica and E. papillata, and obviously representing early ontogenetic stages of these species (Fig. 5a–b). The Sedan material thus provides the basis for the first detailed evaluation of the postlarval development in two extinct ophiuroid taxa, a comparison with extant examples (Stöhr 2005) and its possible phylogenetic implications.
Asteroids
The asteroid assemblage is dominated by (about 70%) Tropidaster pectinatus Forbes, 1850, well known from abundant well-preserved entire specimens from a single occurrence (Pliensbachian of the Mickleham Tunnel, Gloucestershire, UK; Wright 1863 pl. 3; Blake 1996). The taxon has five short, broad arms, and highly distinctive short, broad adambulacral ossicles, which carry a single transverse row of spines. The Sedan material includes entire post-larval individuals (R = 1 mm) (Fig. 5c) and numerous fragments (Fig. 5d) with in situ spines, which allow both a complete ontogeny to be described, and a detailed appraisal of spine morphology and function. In particular, the comb-like adambulacral spines (Fig. 5e, f) are not known in any other asteroid taxon, but show some similarities with certain ophiuroid spines. Abactinal paxillar spines are preserved in life position, cemented by pyrite (Fig. 5g). Additionally, the residues have yielded numerous well-preserved dissociated Tropidaster pectinatus ossicles of all types (Fig. 5h), which permit direct comparison with extant spinulosid taxa (Gale 2010). These ossicles show that Tropidaster is a highly derived taxon with closest affinities to the Solasteridae, and is not related closely to either Korethrasteridae or Pterasteridae (compare Blake 1996).

aInexpectacantha acrobatica Thuy, 2010, articulated young postlarva (GZG.INV.30559), ventral view; scale bar equals 200 μm bE. papillata Thuy, 2010, articulated young postlarva (GZG.INV.30560), ventral view; scale bar equals 500 μm. cTropidaster pectinatus Forbes, 1850, early juvenile (R = 1 mm) (NHM EE 13980), abactinal aspect; note large terminal ossicles and interradial grooves; scale bar equals 500 μm. dTropidaster pectinatus Forbes 1850, arm fragment of small individual (R approximately 15 mm) (NHM EE 13981), actinal view; scale bar equals 1 mm. eTropidaster pectinatus Forbes, 1850, partly dissociated individual (NHM EE 13982) showing adambulacral and comb-like adambulacral spines, actinal view; scale bar equals 500 μm. fTropidaster pectinatus Forbes, 1850, lateral adambulacral spines in situ on adambulacrals of small individual (NHM EE 14020); note articulation surfaces and muscle insertion sites; scale bar equals 100 μm. gTropidaster pectinatus Forbes, 1850, abactinal surface of same specimen, showing clusters of spines still attached to terminations of paxilliform abactinal ossicles, essentially in life position; scale bar equals 100 μm. hTropidaster pectinatus Forbes, 1850, isolated adambulacral ossicle (NHM EE 14021), oblique abactinal view, showing preservation of articular surfaces and muscle insertion sites; scale bar equals 500 μm. All from the Pliensbachian Davoei Zone of Sedan, Ardennes, France; level A, road cutting
Over 30% of the remaining asteroid material comprises arm fragments and ossicles belonging to an unusual, highly derived, multiarmed taxon, close to (but probably not congeneric with) Plumaster ophiuroides Wright, 1863, and here referred to provisionally as Plumaster sp. nov. Plumaster ophiuroides was described on the basis of a single specimen showing a well-preserved actinal surface from the Pliensbachian of the Yorkshire coast, UK (Wright 1863, pl. 5 Fig. 1). Gale (this volume) has established the Plumasteridae to accommodate these asteroids which are superficially similar to multiarmed solasterids, but in morphological detail display numerous unique characters, entirely unknown in any other asteroids (Gale 2010). The adambulacral plates (Fig. 6a, b) are concavo–convex, and successive ossicles articulate by means of a transverse row of interlocking grooves and ridges, similar to those developed on ophiuroid lateral arm plates (Fig. 6c, f). Abactinal ossicles carry single, large, centrally placed bosses for spine articulation (Fig. 6d), with a surrounding (variably well-defined) areole for muscle articulation. These closely parallel the development of echinoid tubercles (Smith 1980). The spines are elongated and composed of glassy trabecular ridges. One arm fragment shows coarsely pyritised tube feet. Small lenses of asteroid debris from Sedan are made up entirely of partially comminuted ossicles of Plumaster sp. nov., and attest to the presence of a predator feeding selectively on this taxon, probably a ray (D.J. Ward, pers. comm.) (Fig. 6e). A notable feature of the preservation of the Plumaster sp. nov. material from Sedan is that the spines are invariably broken between 0.5 and 1.5 mm from the basal attachment. The irregular surfaces of these broken spines attest to their very early date, before decomposition of the stroma and subsequent infilling of the stromal spaces with syntaxial calcite. They almost certainly represent pre-burial abrasion, probably by entrainment in storm waves.
Less than 1% of the remaining asteroid material belongs to other taxa, which are represented by a probable asteriid (circumoral ossicles), an indeterminate paxillosid (circumoral), and two undescribed spinulosids (oral ossicles). The complete dominance of Tropidaster and Plumaster in relatively shallow water clay facies of Pliensbachian age across western Europe (UK, France, Germany, Switzerland) is remarkable.
Holothuroids
At the present stage of investigation, the holothurian fauna is represented by calcareous body wall ossicles only, representing members of two groups: Apodida and Dendrochirotida. The assemblage is dominated by two species with small hooks assignable to the chiridotid subfamily Taeniogyrinae. Achistrum cf. monochordatum Hodson, Harris & Lawson, 1956 is very common, but a detailed SEM study showed remarkably new morphological details, like striae, etc., on the distal part of the hook (Fig. 7i–j). Therefore, there is urgent need of a revision of all Jurassic (and other) holothurian species with hook-like ossicles, as suggested before by Reich (1999, 2010). Unfortunately, there are many Jurassic species, like Achistrum bathonianum Frizzell & Exline, 1956, A. issleri Croneis in Croneis & McCormack, 1932, etc., of which the type material is lost or the whereabouts are unknown.

Plumaster sp., adambulacral ossicles in actinal (a; NHM EE 14022) and abactinal (b; NHM EE 14023) aspects to show exceptional preservation of stereom; scale bars equal 500 μm. Plumaster sp., arm fragment (NHM EE 14024) in actinal (c) and abactinal (d) aspect; note articulating, broken spines. The terminal breaks have irregular surfaces, likely indicating in life or immediately post mortem damage; scale bar equals 500 μm. e Lense composed almost entirely of slightly broken Plumaster ossicles and spines (NHM EE 14025), likely regurgitated by predator, scale bar equals 1 mm. fPlumaster sp., arm fragment (NHM EE 14026) in actinal aspect showing ridge and groove articulation between adjacent adambulacrals, scale bar equals 500 μm. All from the Pliensbachian Davoei Zone of Sedan, Ardennes, France; main echinoderm level, lorry park cutting (a, b) and level A, road cutting (c–f)

Eodiadema sp. a complete test with primary and secondary spines, on the surface of a sideritic nodule (on the same slab than MNHN A30315), lateral view; scale bar equals 1 mm. b Test fragment showing primary and secondary tubercles and spines in place (NHM EE 14027); scale bar equals 500 μm. c Detail of test fragment surface (NHM EE 14027) showing an articulated ophiocephalous pedicellaria with calcitic rod in place; scale bar equals 100 μm. d Detailed view of an ophiocephalous pedicellaria (NHM EE 14027); scale bar equals 100 μm. Eodiadema sp. e Keeled tooth (GZG.INV.45602) in adaxial view; note the aboral end showing zones of secondary plates; scale bar equals 0.5 mm. keeled tooth (GZG.INV.45603) in abaxial view (f), and lateral view (right side) (g); note the aboral end with an outer zone of fine stereom; scale bar equals 0.5 mm. h spectacle ossicle Binoculites terquemi Deflandre-Rigaud, 1952 (GZG.INV.45604); scale bar equals 200 μm. i, j Hook ossicle Achistrum cf. monochordatum Hodson et al. 1956 (GZG.INV.45605); scale bar equals 200 μm. Broken hook ossicle Achistrum sp. nov. (GZG.INV.45606); note the row of spinelets on outer curved edges; scale bar equals 100 μm (k), and 50 μm (l). All from the Pliensbachian Davoei Zone of Sedan, Ardennes, France; level A, road cutting (a) and main echinoderm level, lorry park cutting (b–l)
Surprisingly, two single (broken) hooks (length 0.2–0.3 mm) displayed one short row of medium spinelets on outer curved edges, representing a new chiridotid species (Fig. 7k–l). This new species is comparable to species of Trochodota and Taeniogyrus with similar characteristic spinelets, recently published for the first time by O’Loughlin and VandenSpiegel (2007) within the description of modern material from southern Australia. Momentarily there are no findings of associated taeniogyrinid wheels, probably due to the investigation of only residue >0.1 mm.
Additionally, the sieve residues have yielded a few well-preserved ossicles of Binoculites jurassica Deflandre-Rigaud, 1952 with a length of about 0.3–0.4 mm, representing the first ossicles growth stage (Fig. 7h). According to Gilliland (1992), Binoculites is a dendrochirote sea cucumber, probably of the Cucumariidae, living as an epibenthic suspension feeder. Unfortunately, there is no evidence of pyritised holothurian hard-part or soft-tissue preservation at the time.
Echinoids
Partly dissociated tests and spines of small-sized echinoids are among the most abundant microscopic echinoderm remains from the Sedan Lagerstätte. Intact tests locally occur in great numbers and mostly have their lantern and secondary spines in place while the long, needle-like primary spines are broken at their base. Articulated echinoids with attached intact primary spines are preserved on the surface of the sideritic nodules (Fig. 7a). The echinoids are tentatively assigned to the genus Eodiadema Duncan, 1889, widespread in the Early Jurassic of Europe (e.g., Simms et al. 2004) but in need of taxonomic revision. Thanks to their excellent preservation, the new specimens provide information on poorly known skeletal structures of Eodiadema-like echinoids, including teeth ultrastructure (Fig. 7e–g), apical disc plating and pedicellaria. Exceptionally preserved ophicephalous pedicellariae and spines preserved in place on the surface of the test are illustrated here (Fig. 7b–d). The Sedan material has provided no evidence for any other echinoid taxa.
Preservation
Most of the echinoderm specimens from the Sedan Lagerstätte are preserved as calcitic skeletons, whose fine-scale stereom architecture is, in most cases, hardly blurred by recrystallisation. Even the most delicate structures of comb-like ophiuroid and asteroid appendices or of holothuroid sclerites, for example, are pristinely preserved. Pyritisation occurs in many specimens, especially in the larger articulated ones, preassumably as a consequence of an originally higher concentration of organic matter in these specimens at the time of burial, favouring pyrite formation (Fisher and Hudson 1985). In rare cases, specimens are partly encrusted by coarse grey gypsum, probably formed secondarily as a result of pyrite disintegration in the presence of calcite (Schnitker et al. 1980). The extraordinary preservation of calcitic ultrastructures in the echinoderm skeletons, largely unaffected by recrystallisation, and the preservation of the originally aragonitic, iridescent shell of ammonites, commonly observed in the Sedan Lagerstätte, are both favoured by enclosing argillaceous matrix (Hall and Kennedy 1967).
Nearly all of the articulated echinoderm specimens, including echinoids, from the clays and the surfaces of the sideritic nodules are affected by compaction. Preservation of the original three-dimensional shape in echinoids is only observed in rare, thoroughly pyritised tests from the clays, and in tests inside the sideritic nodules, suggesting that both the nodules and the pyrite formed early during diagenesis.
Soft-tissue preservation has only been observed in a single case of pyritised tube feet in an arm fragment of Plumaster sp. nov. (see above). While, for most echinoderm groups, soft-tissue morphology is of little systematic relevance, the preservation of nonmineralised structures (such as tentacles) is, in contrast, of some importance in holothuroids. Considering the excellent preservation of multisclerite skeletons, requiring rapid and effective burial (e. g., Kerr and Twitchett 2004, Ausich 2001), the reducing conditions in the sediments at least shortly after burial, as indicated by the abundant pyrite, and the presence of syngenetic sideritic nodules, which all favour preservation of nonmineralised structures, it is not unlikely that more traces of soft-tissue, in particular in holothuroids, are among future discoveries of the ongoing sampling and sorting efforts.
Palaeoecology
Initial palaeoecological investigations on the Sedan Lagerstätte suggested that the shelly beds within the clay succession of otherwise low fossil content represented phases of more favourable bottom water conditions and/or of reduced sedimentation rate, allowing for the settlement of Modiolus populations in a near-shore and thus supposedly shallow-water setting on the southeastern margin of the emerged Ardennes landmass (Mouterde et al. 1980, Piénkowski et al. 2008, Thuy 2010). It was furthermore inferred from the co-occurrence of perfectly articulated, partly dissociated and completely isolated skeletal parts that the echinoderms were rapidly and effectively buried as an autochthonous or at least parautochthonous assemblage within the populations of Modiolus bivalves. Ophiuroids with their arms curled in a way suggesting overturning of the disc, as well as common cases of pre-depositional fragmentation among long, multi-element structures, such as crinoid stalks and large ophiuroid arms, were interpreted as causes of strong mechanical stress accompanying burial, most probably under the influence of storm-related currents.
New observations, based on a much larger outcrop area than was accessible at the time of the first investigations, indicate that the Modiolus assemblages were variable in lateral continuity, ranging from small aggregations to continuous beds of considerable lateral extent, as can be inferred from the distribution of Modiolus specimens preserved in live position (Fürsich 1980). Comparisons with recent analogues of the Modiolus assemblages suggest that the preferential occurrence of echinoderms within the bivalve beds might reflect a primary ecological signal. In fact, beds or clumps of extant Modiolus modiolus (Linnaeus, 1758) are known to support significantly more abundant, dense and diverse macrobenthic invertebrate assemblages than surrounding areas devoid of bivalve aggregations by providing stable microhabitats and spatial refuge from predators (Ojeda and Dearborn 1989).
New observations on the Sedan Lagerstätte furthermore document the occurrence of articulated echinoderms in several shelly beds of the succession, which suggest that conditions leading to the formation of the bivalve-echinoderm assemblages and their disruption by major obrution events recurrently prevailed in the depositional area. If the formation of Modiolus beds was triggered by reduced sediment supply and improved bottom water conditions through a decrease in terrigenous discharge, and if burial was mainly caused by storm-related currents, the repeated occurrence of shelly beds with articulated echinoderm assemblages within the succession probably documents climate conditions periodically producing intense storm events occasionally preceded by phases of reduced precipitation rates. Palaeoclimatic reconstructions based on clay mineral composition and oxygen stable isotope, 87Sr/86Sr and Mg/Ca ratios of belemnites suggest relatively uniform and warm seawater temperatures in Peritethyan seas during the Early Pliensbachian Davoei Zone (Rosales et al. 2004) coupled with efficient continental runoff (Dera et al. 2009). Against this large-scale palaeoclimatic background, detailed lithostratigraphical, palaeontological and geochemical analyses of the clay-shelly-bed succession of the Sedan Lagerstätte provide a promising perspective for palaeoclimatic reconstructions at a much higher temporal resolution within the Davoei Zone.
From a community structure perspective, the Sedan Lagerstätte constitutes an interesting and particularly well-preserved example of a habitat, in the present case Modiolus aggregations, closely comparable to extant analogues but with niches mostly occupied by extinct groups.
Further research on the Sedan Lagerstätte and its outstandingly well-preserved echinoderms and other groups will allow a more complete reconstruction of the palaeobiology and palaeoenvironment of this new Lagerstätte.
References
Ausich, W. I. (2001). Echinoderm taphonomy. In M. Jangoux & J.M. Lawrence (Eds.), Echinoderm studies(6 pp. 171–227). Rotterdam: A.A. Balkema.
Blake, D. B. (1996). Redescription and interpretation of the asteroid species Tropidaster pectinatus from the Jurassic of England. Palaeontology, 39(1), 179–188.
Bolette, D. P. (1998). Aspiduriella nom. n. for the genus Aspidura Agassiz, 1835, (Echinodermata: Ophiuroidea: Ophiuridae); preoccupied by Aspidura Wagler, 1830 (Reptilia: Serpentes: Colubridae. Journal of Paleontology, 72(2), 401–402.
Charlesworth, E. (1847). New species of Pentacrinus in the Lias of Yorkshire. The London Geological Journal, 2(96), 131–132.
Croneis, C., & McCormack, J. (1932). Fossil Holothuroidea. Journal of Paleontology, 6(2), 111–148.
de Lamarck, J.-B. P. A. (1799). Prodrome d’une nouvelle classification des coquilles. Mémoires de la Société d’Histoire Naturelle de Paris, 2, 63–91.
Deflandre-Rigaud, M. (1952). Contribution à la systématique des sclérites d’Holothurides fossiles. Bulletin de l’Institut Océanographique, 49(1012), 1–12.
Dera, G., Pellenard, P., Neige, P., Deconinck, J.-F., Pucéat, E., & Dommergues, J.-L. (2009). Distribution of clay minerals in Early Jurassic Peritethyan seas: Palaeoclimatic significance inferred from multiproxy comparisons. Palaeogeography, Palaeoclimatology. Palaeoecology, 271(1–2), 39–51.
Duncan, P. M. (1889). A revision of the genera and great groups of the Echinoidea. Zoological Journal of the Linnean Society, 23, 1–311.
Fisher, I., St, J., & Hudson, J. D. (1985). Pyrite geochemistry and fossil preservation in shales. Philosophical transactions of the Royal Society (B: Biological sciences), 311, 167–169.
Forbes, E. (1844). On the fossil remains of Starfishes of the order Ophiuroidæ, found in Britain. Proceedings of the Geological Society of London, 4, 232–234.
Forbes, E. (1850). Tropidaster pectinatus. Memoirs of the Geological Survey of the United Kingdom, British Organic Remains Decade III. (pp. 1–2) London: Longman, Brown, Green & Longmans.
Frizzell, D. L. & Exline, H. (1956). Monograph of fossil holothurian sclerites. Bulletin of School of Mines and Metallurgy (Technical Series) 89 [1955] (1), 1–204.
Fürsich, F. T. (1980). Preserved life positions of some Jurassic bivalves. Paläontologische Zeitschrift, 54(3/4), 289–300.
Gale, A. S. (2010). The phylogeny of the Neoasteroidea (Asteroidea, Echinodermata). Special papers in palaeontology. (in press).
Gilliland, P. M. (1992). Holothurians in the Blue Lias of southern Britain. Palaeontology, 35(1), 159–210.
Goldring, R. & Stephenson, D. G. (1972). The depositional environment of three starfish beds [Neues Jahrbuch für Geologie und Paläontologie]. Monatshefte (10), 611–624.
Hall, A., & Kennedy, W. J. (1967). Aragonite in fossils. Proceedings of the Royal Society of London (B: Biological Sciences), 168, 377–412.
Heer, O. (1865). Die Urwelt der Schweiz. Zürich: Schultheiß, 1–622.
Hess, H. (1964). Die Ophiuren des englischen Jura. Eclogae Geologicae Helvetiae, 57(2), 756–801.
Hess, H. (2010). Myzostome deformation on arms of the Early Jurassic crinoid Balanocrinus gracilis (Charlesworth). Journal of Paleontology, 84(6), 1031–1034.
Hess, H., & Meyer, Ch A. (2008). A new ophiuroid (Geocoma schoentalensis sp. nov.) from the Middle Jurassic of north-western Switzerland and remarks on the family Aplocomidae HESS, 1965. Swiss Journal of Geosciences, 101, 29–40.
Hodson, F., Harris, B., & Lawson, L. (1956). Holothurian spicules from the Oxford clay of redcliff, near weymouth (Dorset). Geological Magazine, 93(4), 336–344.
Hyatt, A. (1867). The fossil cephalopods of the museum of comparative Zoology. Bulletin of the Museum of Comparative Zoology, Harvard University, 1, 71–102.
Kerr, T. J. V., & Twitchett, R. J. (2004). Experimental decay and disintegration of Ophiura texturata: implications for the fossil record of ophiuroids. In T. Heinzeller & J. H. Nebelsick (Eds.), Echinoderms München (pp. 439–446). London: Taylor & Francis.
Kutscher, M., & Jagt, J. W. M. (2000). Early Maastrichtian ophiuroids from Rügen (northeast Germany) and Møn (Denmark). In J. W. M. Jagt (Ed.), Late Cretaceous- Early Palaeogene echinoderms and the K/T boundary in the southeast Netherlands and northeast Belgium- Part 3 (Vol. 121, pp. 45–107). Ophiuroids: Scripta geologica.
Linnaeus, C. (1758). Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I., editio decima, reformata. Holmiae (Laurentii Salvii), 1–4, 1–824.
Lyman, T. (1869). Preliminary report on the Ophiuridea and Astrophytidae dredged in deep water between Cuba and the Florida Reef, by L.F. de Pourtales, Assist. U.S. Coast Survey. Bulletin of the Museum of Comparative Zoology, Harvard University, 1, 309–354.
Madsen, F. J. (1983). A review of the Ophioleucinae stat. rev. (Echinodermata, Ophiuroidea) with the erection of a new genus, Ophiostriatus. Steenstrupia, 9(2), 29–69.
Martynov, A. (2010). Structure of the arm spine articulation ridges as a basis for taxonomy of Ophiuroidea (a preliminary report). In L. G. Harris, A. S. Böttger, C. W. Walker & M. P.Lesser (Eds.), Echinoderms: Durham. Proceedings of the 12th International Echinoderm Conference, Durham, New Hampshire, USA, (pp. 233–239) 7–11 August 2006. London: Taylor & Francis Group.
Mouterde, R., Tintant, H., Allouc, J., Gabilly, J., Hanzo, M., Lefavrais, A., et al. (1980). Lias. In C. Mégnien (Ed.), Synthèse Géologique du Bassin de Paris (Volume I ed., Vol. 101, pp. 75–123). Stratigraphie et Paléogéographie: Mémoire du B.G.R.M.
O’Loughlin, P. M., & VandenSpiegel, D. (2007). New apodid species from southern Australia (Echinodermata: Holothuroidea: Apodida). Memoirs of Museum Victoria, 64, 53–70.
Ojeda, F. P., & Dearborn, J. H. (1989). Community structure of macroinvertebrates inhabiting the rocky subtidal zone in the Gulf of Maine: seasonal and bathymetric distribution. Marine Ecology Progress Series, 57, 147–161.
Phillips, J. (1829) Illustrations of the Geology of Yorkshire. Part 1. The Yorkshire Coast. (pp. 1–184) London: John Murray.
Piénkowski, G., Schudack, M. E., Bosák, P., Enay, R., Feldman-Olszewska, A., Golonka, J., et al. (2008). Jurassic. In T. McCann (Ed.), The geology of Central Europe, Volume 2: Mesozoic and Cenozoic (pp. 823–922). London: The Geological Society.
Reich, M. (1999). Über Achistrum huckei (Frizzell & Exline 1956) [Holothuroidea, Echinodermata] aus der Unterkreide von Bartin (Hinterpommern). Greifswalder Geowissenschaftliche Beiträge, 6, 383–391.
Reich, M. (2010). The early evolution and diversification of holothurians (Echinozoa). In L. G. Harris, A. S. Böttger, C. W. Walker & M. P. Lesser, (Eds.) Echinoderms: Durham. Proceedings of the 12th International Echinoderm Conference, Durham, New Hampshire, USA (pp. 55–59) 7–11 August 2006. London: Taylor & Francis Group.
Rosales, I., Quesada, S., & Robles, S. (2004). Paleotemperature variations of Early Jurassic seawater recorded in geochemical trends of belemites from the Basque-Cantabrian basin, northern Spain. Palaeogeography, Palaeoclimatology, Palaeoecology, 203, 253–275.
Schnitker, D., Mayer, L. M., & Noirton, S. (1980). Loss of calcareous microfossils from sediments through gypsum formation. Marine Geology, 36(3–4), M35–M44.
Seilacher, A., Reif, W.-E., & Westphal, F. (1985). Sedimentological, ecological and temporal patterns of fossil Lagerstätten. Philosophical Transactions of the Royal Society of London (B: Biological Sciences), 311, 5–23.
Simms, M. J., Chidlaw, N., Morton, N. & Page, K.N. (2004). British Lower Jurassic stratigraphy. Geological conservation review series, No 30 (pp. 1–458) Peterborough: Joint Nature Conservation Committee.
Smith, A. B. (1980). The structure and arrangement of echinoid tubercles. Philosophical Transactions of the Royal Society (B: Biological Sciences), 289, 1–54.
Smith, A. B., Paterson, G. L. J., & Lafay, B. (1995). Ophiuroid phylogeny and higher taxonomy: morphological, molecular and palaeontological perspectives. Zoological Journal of the Linnean Society, 114, 213–243.
Sowerby, J. (1822). The mineral conchology of Great Britain. Meredith, London, 1–4, 1–383.
Stöhr, S. (2005). Who’s who among baby brittle stars (Echinodermata, Ophiuroidea): Postmetamorphic development of some North Atlantic forms. Zoological Journal of the Linnean Society, 143, 543–576.
Thuy, B. (2010). Exceptionally well preserved brittle stars from the Pliensbachian (Early Jurassic) of the French Ardennes. Palaeontology 54(1) (in press).
Wright, T. (1863). A Monograph on the British fossil Echinodermata from the Oolitic Formations. Volume second. Part 1. On the Asteroidea. The Palaeontographical Society 15 [for 1861], 1–130.
Wright, T. (1866). A Monograph on the British fossil Echinodermata from the Oolitic Formations. Volume second. Part 2. On the Ophiuroidea. The Palaeontographical Society 18 [for 1864], 132–154.
Acknowledgments
We would like to express our thanks to Hans Hess (Basel, CH) and Christian Meyer (Natural History Museum Basel, CH) whose comments greatly improved this manuscript, and to Tanja Stegemann (University of Göttingen, D) for assistance during field work.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Thuy, B., Gale, A.S. & Reich, M. A new echinoderm Lagerstätte from the Pliensbachian (Early Jurassic) of the French Ardennes. Swiss J Palaeontol 130, 173–185 (2011). https://doi.org/10.1007/s13358-010-0015-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-010-0015-y