
Abstract
The asteroid fauna from the Late Oxfordian marls (bifurcatus Zone, stenocycloides Subzone) of Savigna includes 11 taxa, distributed between 8 families, of which 1 (Plumasteridae) is new. A goniasterid of distinctive morphology, Hessaster longimarginalis, is described as new, as is an asteriid, Savignasterias villieri. The material was recovered by surface picking and processing over 1,000 kg of sediment, and includes 28 partial individuals and approximately 2,000 isolated ossicles in excellent preservation, which show fine details of the stereom architecture and additionally allow ontogenetic changes to be described. Detailed comparison with extant asteroids enables diverse ossicle types to be assigned precisely to individual taxa. The phylogenetic relationships of individual fossil species with living taxa are determined using characters derived from numerous skeletal elements, and it is shown that the benthopectinid and pterasterid species present in the Savigna fauna are basal to their respective families. In comparison with other assemblages collected from Jurassic clay facies, the Savigna asteroid fauna is unusually diverse. Importantly, the fauna includes elements typical of present day deep sea environments (bathyal and abyssal), living abundantly in Oxfordian shelf seas of about 50 m depth.
Similar content being viewed by others


Introduction
Much attention has been focused upon the rare finds of articulated, beautifully preserved asteroid specimens. Such material is very important in our understanding of ancient faunas, as well demonstrated by Hans Hess’ thorough description and illustration of the Schinznach fauna from the Hauptrogenstein (Bajocian) of Switzerland (Hess 1972) which documented the taxonomy and diversity of a Jurassic shallow marine echinoderm assemblage. However, it is unwise to rely entirely upon the infrequently found entire specimens, for two reasons. Firstly, they often reveal little, if any, detail of the internal skeletal structures of the taxonomically important ambulacral groove and mouth frame ossicles, and secondly, they only represent a fraction of the real diversity of ancient asteroid assemblages.
Although dissociated asteroid ossicles have been collected and described for a long time (e.g. Desmoulins 1832; Goldfuss 1831), study has mostly concentrated upon the large and conspicuous marginal ossicles of astropectinids and goniasterids, and the enlarged abactinal and marginal ossicles of families such as stauranderasterids and sphaerasterids which can be readily collected in the field. Our knowledge of the fossil record of these families is consequently relatively good, at least insofar as many genera and species have been described. However, many asteroid taxa are themselves small, and consequently have very small ossicles, invisible to field collectors. Processing bulk sediment to concentrate residues, and picking asteroid ossicles from these residues, can provide very useful information on the diversity and occurrence of poorly known families, such as the Late Cretaceous pterasterids (Villier et al. 2004a).
Asteroid skeletons are complex structures, including 11 major constructional skeletal elements, and diverse spine morphologies (Turner and Dearborne 1972). Consequently, the ability to identify diverse ossicle types in the fossil state and compare these with extant families requires a detailed comparative knowledge of the skeletal anatomy of extant asteroid families. This work was pioneered by Blake (1973), and continued using the SEM by Gale (in press). The asteroid skeleton, with its diverse and morphologically complex elements, is in some ways comparable with the vertebrate skeleton. It is therefore possible, with care, to virtually reconstruct taxa from isolated ossicles of different types in exactly the same way as, say, a Pleistocene mammal worker can confidently identify an extinct species from a diversity of skeletal elements. It is always necessary to be cautious, especially when a number of closely related species are present in an assemblage.
The description of asteroids from multiple skeletal elements has a number of advantages. Firstly, the amount of morphological information that can be obtained from a suite of ossicles is considerable, and allows very detailed comparison with extant forms, permitting the reconstruction of phylogenies. In this way, Gale (in press) was able to demonstrate that Oxfordian taxa based on ossicles, and related to extant benthopectinids and pterasterids, lack critical skeletal characters present in all representatives of the living families. They, therefore, represent basal members of their respective groups. In turn, these features are directly related to specific life habits, and it can thus be demonstrated, e.g., that Jurassic pterasterids did not possess an abactinal canopy, and that benthopectinids lacked longitudinal muscles in the arms (Gale in press).
It has become evident over the past few years that marine clay deposits often contain well-preserved asteroid ossicles, and interest has been focused upon Jurassic clays. The most abundant and diverse fauna yet found is from the upper Oxfordian of Savigna, in the French Jura, originally discovered by Hans Hess (1966). With the encouragement of Hans, I revisited the section in 2007, and have subsequently collected and processed approximately 1 ton of material. This has yielded over 2,000 asteroid ossicles, belonging to 12 taxa, 4 of which were described by Gale (in press). In this paper, I describe the remaining taxa and review the entire fauna from both phylogenetic and palaeoecological perspectives.
Locality, stratigraphy and palaeoenvironment
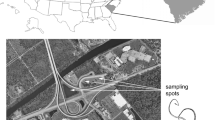
The village of Savigna is situated approximately 10 km SSW of the town of Orgelet in the Départment of Jura in eastern France (Fig. 1a). Approximately 500 m southwest of the church, 100 m to the north of the road to Fétigny, a series of natural exposures of Oxfordian calcareous clays and marly limestones belonging to the lower Couches d’Effingen (Enay 1966) are exposed in the banks of a small stream (Fig. 1a, locality 2a: N 46°26′09.7″, E 005°34′55.1″, 2b: N 46°26′11.3″, E 005°34′58.2″). The succession is continued downwards in a shallow excavation 50 m to the southwest, immediately west of the Fétigny road (Fig. 1a, locality 1: N 46°26′10.3″, E 005°35′04.8″). The two successions can be correlated to make a composite outcrop of nearly 15 m, comprising three thin marly limestones and intervening calcareous clays (Fig. 2). The Savigna exposures are assigned to the stenocycloides Subzone of the bifurcatus Zone, Late Oxfordian, dated to approximately 157–158 Ma.

Location of Savigna section. a Present geography, b palaeogeographical map to show position of Jura Platform in the Oxfordian. After Védrine and Strasser (2009)

Composite stratigraphical section at Savigna, combining succession at localities 1 and 2 (Fig. 1a), showing distribution of commoner asteroid taxa
The weathered surfaces of the clays are highly fossiliferous, with abundant columnal fragments of the crinoid Balanocrinus, zeillerid and acanthothyrid brachiopods, serpulids, pyritised nuclei of ammonites and diverse calcitic bivalves. The clay grade of the sediment, total bioturbation, and lack of accumulations of coarser debris suggest deposition below storm wave base (in excess of 50 m; see Sahagian et al. 1996). The abundance of suspension feeding crinoids, bivalves, serpulids and brachipods would support a shallower rather than deeper estimate, around 40–60 m. The high diversity and density of benthos indicate an oxygenated palaeoenvironment, with reasonable productivity. During the Oxfordian, the Jura was situated on a platform adjacent to the Helvetic Basin, on the north side of the Tethys spreading ridge (Védrine and Strasser 2009; Fig. 1b herein).
Materials and methods
Calcareous clay was collected from a series of fossiliferous levels at Savigna, and dried in the sun. It was processed in a clay machine (Ward 1981) using a 0.25 mm sieve, and the residue was dried, graded and picked and sorted into ossicle types and taxa. Ossicles were cleaned using an ultrasonic tank, and mounted for SEM imaging. Ossicles were identified and assigned to taxa with reference to material of extant species (see Gale in press). For abbreviations, see Table 1. Museum abbreviations: BMNH, Natural History Museum, London; CAMSM, Sedgwick Museum, Cambridge, UK; Natural History Museum Basel, Switzerland, NMB. Abbreviations for morphological features follow Gale (in press) and are included in Table 1.
Systematic palaeontology
Paxillosida PERRIER, 1884
Astropectinidae GRAY, 1840
Pentasteria VALETTE, 1929b
Pentasteria (Pentasteria) longispina HESS, 1968

Pentasteria (P.) longispina Hess ossicles, Oxfordian, stenocycloides Subzone, Savigna, 2b horizon (a, b, e, g, h) and Red Nodule Bed, Oxford Clay, costicardia Subzone, Weymouth, Dorset, UK (c, d, f). Adambulacral ossicles (BMNH EE 13962), enlargement of abradial abactinal surface to show saadm (a); median adambulacral (EE 13961) in abactinal (b) and actinal (a) views. Ambulacral ossicle in actinal (c) and distal (d) views (EE 13934). Oral ossicle in interradial (f1) and radial (f2) views (EE 13937). IM ossicles of juvenile in actinal (g1, h) and proximal (g2) views (g EE 13963, h EE 13964). a ×20; rest ×15

Drawings of P. (P.) longispina ossicles; a, b, d, e are from the costicardia Subzone Red Nodule Bed, Oxford Clay, Weymouth Dorset, UK; c is from horizon 2b, stenocycloides Subzone, at Savigna. Bases of ambulacral ossicles to show adamb articulations and muscle sites, in oblique distal (a) and oblique proximal (b) views (BMNH EE 13933-4). Adambulacral ossicle, in actinal (c1) and abactinal (c2) views (BMNH EE 13935). Circumoral ossicle in abactinal (d1) and actinal (d2) views (BMNH EE 13936). Oral ossicle in interradial (e1) and radial (e2) views (BMNH EE 13937). Scale bar 5 mm
1968 Pentasteria (Pentasteria) longispina Hess, 607–614, figs. 1–3, pl. 1.
1975 Pentasteria (Pentasteria) longispina Hess, pl. 1, pl. 8, figs. 7, 8.
1999 Pentasteria (Pentasteria) longispina Hess, figs. 2, 3.
Type:
Specimen figured by Hess (1968), NMB M 8748, from the Late Oxfordian (bifurcatus Zone) of Schöfgraben, Weissenstein. The species is well known from a slab including 12 superbly preserved individuals, from Schöfgraben, Weissenstein, figured by Hess (1999), fig. 2 (NMB M 17416).
Referred material:
The Savigna material comprises numerous (>150) well-preserved isolated ossicles including ambulacrals, adambulacrals, orals, circumorals and marginals (figured material BMNH EE 13935, EE 13961-4). Additional material from the Early Oxfordian Red Nodule Bed (costicardia Subzone) of Weymouth, Dorset, is also figured (BMNH EE 13933-4, 13936-7).
Description:
P. (P.) longispina is well known from entire individuals and marginal ossicles, and the overall morphology has been thoroughly described. The arms are long and some superomarginals carry tall conical spines, and the disc is small with acute interradii (Hess 1999). The morphology of isolated ambulacral groove and mouth frame ossicles has never been hitherto described for any species of Pentasteria.
In actinal view, the proximal adambulacrals (Figs. 3b, e, 4c) are transversely rectangular, become square in the mid-radius, and are slightly elongated distally. Padam and dadam surfaces are transversely broad and short, subparallel, and padam is borne on a raised platform. Ada2 forms a prominent boss immediately distal and abradial to the adpm, and the adad articulation is flush with the abactinal surface, set on a short proximal protuberance. Ada3 is a flat, oval surface adjacent to the dadam, and ada1 is transversely broad, short and concave. The attachment for the superambulacral muscle (Fig. 3a, saadm; see Heddle 1967, fig. 6) is positioned on a short process on the abradial margin between the padam and dadam, and is very sharply defined on the proximal adambs of larger specimens. The actinal face is divided into a proximal, raised, transverse spine bearing surface and a concave distal surface for articulation with the distal adamb. A single, centrally placed base for a large subadambulacral spine is surrounded by an irregular scatter of small spine bases. A well-defined, curved row of up to 10 fs bases is present on the actinal adradial margin.
The ambulacrals (Fig. 3c, d) are short and broad, and the amb base is short, parallel sided and broad (over 30% of the transverse dimension). The wings for padam and dadam are short and broad. On the proximal side, ada1 forms a very broad, short strip, and discrete, oval ada2 and ada3 are present on the distal side.
The body of the oral ossicles in P. (P.) longispina (Figs. 3f1, f2, 4e1, e2) is oval and moderately elongated and the apophyse is tall and prominent with a prominent flange for insertion of the riom. The contact surface for the first adamb (oradm, orada) is large. The proximal blade is short and blunt, and a flattened facet for articulation with the adjacent oral across the radius is present (rart). Deep notches for articulation of the 8–10 oral spines are present. The three centrally placed suboral spine bases are large and u-shaped, and surrounded by smaller spine bases. On the interradial face, iioa structures are present but dentition is absent.
Marginals of presumed juvenile individuals of P. (P.) longispina are common in the Savigna residues (Fig. 3g, h). The ims are rectangular–trapezoidal in actinal view, with a poorly defined intermarginal ridge, and the external face carries even sized spine attachment sites. These ossicles resemble the species described as P.? liasica by Villier et al. (2004b).
Discussion:
The first description of the morphology of ambulacral and mouth frame ossicles of a Jurassic astropectinid permits detailed comparison to be made with extant members of the family. This is important, because Blake (1986) transferred a typical member of Pentasteria (Pentasteria) to the goniasterid genus Pseudarchaster on the basis of marginal morphology. Gale (in press) argued that P. (P.) portlandensis Hess 1955, rather than being a Pseudarchaster, is a true astropectinid on the basis of marginal spine morphology and arrangement. The ambulacral and adambulacral morphology of P. (P.) longispina described here is typically astropectinid in the overall shape of the ossicles and the arrangement of the ambulacral–adambulacral contact structures (e.g. single, transversely broad, short ada1, knob-like ada2, prominent adada flush with abactinal surface of ossicle). It also possesses a facet for insertion of the superambulacral muscle (saadm), only found paxillosids. However, there are significant differences in the morphology of ambulacral groove structures between P. (P.) longispina and extant astropectinids. In many astropectinids (e.g. Astropecten, Ctenopleura, Craspidaster, Ctenophoraster), padam and dadam are positioned in a transverse arrangement on the adamb, corresponding to the strong asymmetry of the ambulacral base (padam on a narrow proximal wing of the amb base, dadam on the broader proximal wing; see Gale in press, text-fig. 10C, pl. 6, figs. 10, 11). The orals of extant astropectinids have a rather uniform morphology, with a low apophyse approximately flush with the distal abactinal margin of the oral (see Gale in press, text-fig. 16A; Blake 1973, fig. 1N, O), strong dentition, a short proximal blade, and an external surface which carries irregularly sized and arranged spine bases. Additionally, the posterior radial surface which contacts the first adamb is large and oval. The oral of P. (P.) longispina has a tall apophyse and lacks dentition, but otherwise is similar to astropectinids in construction. In summary, the detailed skeletal morphology of P. (P.) longispina supports its position as a basal astropectinid.
Cribellina FISHER, 1911
Goniopectinidae VERRILL, 1899
Chrispaulia GALE, 2005
Chrispaulia jurassica GALE, in press
Fig. 5a–f

Chrispaulia jurassica Gale in press. Marginal ossicles, Savigna 1 horizon. a Median SM in lateral aspect (BMNH EE 13938). b SM from mid-arm, lateral aspect (BMNH EE 13591). c Distal SM, lateral aspect (BMNH EE 13585). IM from base of arm (BMNH EE 13590), lateral (d1) and proximal (d2) views. e Distal IM, lateral aspect (BMNH EE 13587). All ×20
2010 Chrispaulia jurassica Gale, in press, pl. 18, figs. 1–11.
Type material:
The holotype is a superomarginal ossicle (BMNH EE 13586). A suite of marginal ossicles are paratypes (EE 13585-91).
Referred material:
72 marginal ossicles from Savigna, most from the S1 fauna (BMNH EE 13938; EE 13585-91).
Description:
The interradial sm (Fig. 5a) are nearly square in lateral aspect, narrow, lacking an abactinal face, and the external faces are gently convex. The majority of the lateral face is occupied by even, fine rugosities which bore fasciolar spines. Distally, the sm become blocky, more elongated, and have well-demarcated lateral and abactinal faces (Fig. 5b, c). The fasciolar surfaces form narrow strips along the proximal and distal margins, and a central area of coarse rugosities is present. The rugosities cojoin irregularly to form transverse strips on the lateral faces. The actinal margin of the im (Fig. 5d–f) is slightly concave, and occupied by coarse, evenly sized rugosities. The lateral face of the distal im carries one to three large, bifid spine bases arranged diagonally (Fig. 5d).
Discussion:
Extant goniopectinids, and the Cretaceous species Chrispaulia radiata, are characterised by the presence of articular notches for lamellar cover spines on the proximal and distal margins of the raised central regions of the marginals (Gale 2005). These are lacking in C. jurassica. However, the goniopectinid affinities of a species close to C. jurassica are confirmed by an undescribed species of Chrispaulia from the Hauterivian Tealby Clay of Yorkshire, UK, and St Pierre de Chevennes, Isère, France, which includes oral and circumoral ossicles typical of extant Goniopectinidae, and closely comparable with those of the present day genus Goniopecten.
Benthopectinidae VERRILL, 1899
Jurapecten GALE, in press
Jurapecten hessi GALE, in press
Fig. 6a–h

Jurapecten hessi Gale in press. Savigna, 2a horizon. Ambulacral ossicles in actinal (a) and distal (b) views (BMNH EE 13595, EE 13597). c Distal sm (BMNH EE 13602), and d distal im in lateral views (BMNH EE 13600). e Oral in radial view (BMNH EE 13606). Adambulacral ossicle in actinal (f) and abactinal (g) views (BMNH EE 13939, EE 13599). h Terminal ossicle in abactinal view (BMNH EE 13609). See Gale (in press, pls. 18, 19) for more detailed description and illustration. All ×20
2010 Jurapecten hessi Gale, pl. 19, figs. 1–6, 8–12; pl. 20, figs. 5–8, 10, 12, 14, 16, 18.
Type material:
An individual showing ambulacrals and adambulacrals is holotype (BMNH EE 13594). The paratypes are isolated ossicles (EE 13592-13609).
Referred material:
Five individuals, comprising associated but disarticulated ossicles, and approximately 300 isolated ossicles.
Discussion:
The species was described and illustrated by Gale (in press), and details of each ossicle type (ambulacrals, adambulacrals, orals, circumorals, terminals, marginals) compared in detail with those of extant benthopectinids. The highly specialised ambulacral–adambulacral articulation (see Blake 1973) confirms the benthopectinid affinities (Fig. 6a), but the absence of transverse ridges on the abactinal surfaces of the ambulacrals of J. hessi demonstrates that the species must have lacked the longitudinal arm muscles present in all extant members of the family. The species was very small (R < 15 mm), and is now known also from the Callovian lamberti Zone of Round House Farm, near Swindon, UK.
Valvatida PERRIER, 1884
Goniasteridae FORBES, 1841
Tylasteria VALETTE, 1929b
Tylasteria jurensis (MÜNSTER in GOLDFUSS, 1831)

Tylasteria jurensis (Münster in Goldfuss). Arm fragment (a) and ossicles probably originating in same individual, horizon 2b, Savigna BMNH EE 13940). a Abactinal aspect of distal arm, to show abactinal ossicles and sm spination. b Same, in actinal aspect, to show im spination. c Isolated distal IM. b Ambulacral ossicle in actinal aspect. Adambulacral ossicles in actinal (d) and actinal (g) views; compare with Fig. 6d. e Abactinal ossicle in oblique view. f Spine from abactinolateral margin of sm (compare a). h Actinal ossicle in actinal aspect. a1 ×4, b ×6, c, d, g ×10, e, f, h ×20

Drawings of Tylasteria jurensis ossicles. Savigna 2b horizon, stenocycloides Subzone. Material associated with the individual figured in Fig. 7 (BMNH EE 13940). Circumoral ossicle, in a1 abactinal and a2 actinal aspects. Oral ossicle, in interradial (b1) and radial (b2) aspects. c Ambulacral ossicle, base in oblique proximal aspect to show articulation (ada1) and muscle (padam) sites. Adambulacral ossicle in distal (d1) and abactinal (d2) aspects. Scale bar 5 mm
p1831 Asterias jurensis Münster in Goldfuss, pl. 63, fig. 6a, b only.
p1886 Asterias impressae Quenstedt, p. 583, pl. 73, figs. 60–80.
1928 Tylaster priscus, Valette, p. 62, fig. 6.
1929 Tylaster priscus Valette, p. 29, pl. 5, figs. 1–5.
1975 Tylasteria sp. Hess, p. 35, pl, 8, fig. 18.
1992 Tylasteria prisca (Valette), Bréton, pp. 252–259, pl. 28, fig. 11, pl. 29,
pl. 30, figs. 1–3.
Types:
The arm fragment figured by Münster in Goldfuss from Würtemburg (1831, pl. 63, fig. 6a, b), probably of Oxfordian age, is selected as lectotype. Whereabouts unknown, possibly either Berlin or Munich collections. Goldfuss’ figured material referred to A. jurensis comprises a goniasterid arm fragment (pl. 63, fig. 6a), of which a single marginal ossicle is enlarged (pl. 63, fig. 6b). The excellent quality of the illustrations enables identification of the fragment as a Tylasteria conspecific with material from the Oxfordian of France, as shown by the presence of enlarged conical spines on the abactinolateral margin of the sm. However, the remaining items included in Fig. 8 of Münster in Goldfuss (1831, pl. 63) are a median superomarginal of the goniasterid Metopaster uncatus (Forbes 1848; Goldfuss 1831, pl. 63, fig. c–e), from Late Cretaceous Chalk (Turonian–Campanian; see Gale 1987) and three abactinals of a distinctive stauranderasterid (Goldfuss 1831, pl. 63, figs. f–h), presumably of Late Jurassic age.
Referred material:
A partial individual from the horizon 2b at Savigna includes a well-preserved arm fragment with superficial spines in situ (BMNH EE 13940). Over 100 ossicles, including marginals, abactinals, actinals, orals, adambulacrals and ambulacrals from the same horizon, some of which originate from the same individual.
Description:
A large species, in which R exceeds 10 cm. The disc is broad, the interradii gently curved, and the arms taper slowly. The interradial marginals are proportionately tall and short, and their lateral surfaces are vertical or slightly overhanging, forming a wall-like border. The external surfaces of the marginals (Fig. 7a) have a honeycomb-like sculpture of polygonal granule pits which decrease in size at the borders. The in situ spines are pentagonal and hexagonal in outline, and form a closely packed array of low domes (Fig. 7c). Single large conical spines are present on the abactinolateral margin of some superomarginals; these carry rows of short thorn-like spines (Fig. 7f). The abactinal ossicles are small and have a paxilliform morphology (Fig. 7e). The actinals are flat and rectangular in outline, and imbricate (Fig. 7h).
The adambulacrals are cuboidal (Figs. 7d, g, 8d1, d2), and the convex actinal surface which bears two longitudinal ridges for attachment of the subadambulacral spines. An adradial ridge carries 8–10 short, transverse, slit-like attachment sites for the furrow spines. An asymmetrical adpm is present. On the abactinal surface, articulation surfaces ada2 and ada3 are large and flat and ada2 is positioned on the abradial side of the dadam. Ada1a is made up of one or two concave facets, and ada1b is directed distally. The double ada1a developed on some ossicles is unique among asteroids (compare Gale in press, text-fig. 10). The ambulacrals (Fig. 7b) are stout, with a short, narrow waist, and articulation surfaces (lia) on the ambulacral heads are set on discrete processes. The amb base is rounded in actinal aspect, with prominent ada1 articulation surfaces (Fig. 8c).
The oral body (Fig. 8b1, b2) is elongated, and the proximal blade prominent and acutely terminated; a flat surface (rart) articulates radially with the adjacent oral. The oral-first adamb contact surface is oval and occupies the distal portion of the radial face. The triangular actinal face carries 15 notches for oral spines, and 3–4 rows of attachment bases for suboral spines. The apophyse is tall and stout, with a prominent flange for insertion of the riom. The interradial face shows two to three ioa surfaces, and an elongated site for insertion of aciim. The circumorals are sturdy and carry strong dentition (Fig. 8a1, a2).
Discussion:
This material from Savigna provides the first detailed information on the amb groove and mouth frame ossicle morphology of a Jurassic goniasterid. Although the overall morphology of the ossicles is similar to that of extant species of goniasterid (Gale in press), the amb–adamb contact (ada1a with double surface) is highly distinctive and distinguishes Tylasteria from other goniasterid genera.
Noviaster VALETTE, 1929a
Noviaster sp.
Fig. 9g

Hessaster longimarginalis gen. et sp. nov. (a–i), Noviaster sp. (g) and undescribed goniasterid genus (j), all from Savigna horizon 2b. Median unpaired IM of immature individual, in abactinal (a) and actinal (e) views (BMNH EE 13941-2). Abactinal ossicles in abactinal (b, d), lateral (f, h) and internal (c) aspects (c, h BMNH EE 13943, b EE 13944, d EE 13945, f EE 13946). i Actinal ossicle in actinal view (EE 13949). g Marginal of Noviaster sp. (either SM or IM) in actinal or abactinal aspect (BMNH EE 13947). j Marginal (SM or IM) of undescribed goniasterid in actinal or abactinal aspect (BMNH EE 13948). ×25
Referred material:
Three isolated marginal ossicles, of which one is figured (BMNH EE 13947).
Description:
Small marginal ossicles, which possess a depressed narrow rim on the external face and a sculpture of rather coarse, deeply impressed spine pits (Fig. 9g). The marginals imbricated strongly proximally, and proximal and distal surfaces are slanted at about 30° to the long axis of the arm. Discrete abactinal and lateral faces are present.
Discussion:
The material is referred provisionally to Noviaster on account of the distinctively imbricated marginals, also seen on specimens of N. polyplax (Hess 1972, pl. 1, figs. 3–5).
Undescribed goniasterid (?) genus
Fig. 9j
Referred material:
Four marginal ossicles, one figured (BMNH EE 13948).
Description:
Marginal ossicles (Fig. 9j) elongated, low, rhombic in abactinal–actinal aspect. Marginals imbricated strongly proximally, and proximal and distal surfaces are angled at 45° to long axis of the arm. Central region of external face with sculpture of short, coarse, anastomosing ridges.
Discussion:
These ossicles are very elongated marginals, with slanted proximal and distal articulation surfaces similar to those of Noviaster (fig. 7g) and an unusual sculpture. They probably belong to an undescribed goniasterid of unusual morphology.
Genus Hessaster nov.
Derivation of name:
The genus is named in honour of the exceptional contributions of Hans Hess to the study of fossil echinoderms.
Diagnosis:
A goniasterid which possesses elongated, unpaired, pentagonal interradial marginals, elongated, narrow, second and third marginals, and distally progressively shorter marginals. Superomarginals smooth, inferomarginals with sculpture of coarse rugosites and intervening granule pits.
Type species:
Hessaster longimarginalis sp. nov, Oxfordian, French Jura. No other species referred to the genus.
Hessaster longimarginalis gen et sp. nov.

Hessaster longimarginalis gen. et sp. nov. Isolated ossicles, Savigna 2a horizon. a Holotype (BMNH EE 13965), unpaired median sm in abactinal aspect. Unpaired median ims in actinal (b, c) and abactinal (d) views (EE 13966-68). g sm in abactinal view (EE 13969). e, f, j ims in actinal aspect (EE 13970-72). Adambulacrals in actinal (h) and abactinal (k) aspects (EE 13973-74). i Primary (?) abactinal in abactinal aspect (EE 13975). Oral ossicles in interradial (l) and radial (m) aspects (EE 13976-7). All ×15

Reconstruction (a) and median interradial marginal profiles (b, c) of Hessaster longimarginalis gen et sp. nov. Scale bar for a 10 mm
Types:
The unpaired interradial SM (Fig. 10a; BMNH EE 13965) is holotype, the other figured ossicles are paratypes (BMNH EE 13941-46; EE 13966-77).
Referred material:
60 marginal ossicles, 10 adambulacrals, 15 ambulacrals, 8 oral ossicles, 2 actinals, 16 abactinals. Late Oxfordian stenocycloides Subzone of Savigna.
Description:
The unpaired interradial marginal ossicles (Fig. 10a–d) are pentagonal in actinal/abactinal view, and slightly less than twice as long as broad. The lateral margins are gently concave, and the broadest part of the ossicle is coincident with the interradius. The lateral and abactinal/actinal faces are set at right angles and the lateral margin is evenly rounded (Fig. 11b, c). First sm and im ossicles articulate by a long, narrow flat facet with a small, internally directed, central spur (Fig. 10d). The largest unpaired marginal ossicles are 3 mm in length. The second marginal pair (Fig. 10d, g) are rectangular, twice as long as broad and equal in length to the first marginals. The internal and lateral margins are slightly curved. The third marginal pair (Fig. 10f) are rectangular, longer than broad, and taper slightly distally. More distal marginals (Fig. 10e) become progressively shorter, and it can be estimated that a total of six or seven pairs were present in each side of the arm distal to the unpaired interradial marginals (Fig. 11a). The superomarginals have a relatively narrow abactinal face, and a crimped inner margin (Fig. 10a). The surface is smooth or slightly pitted. The inferomarginals have broad, slightly concave actinal surfaces with a sculpture of coarse, irregular rugosities (Fig. 10b–d).
Abactinal ossicles are provisionally assigned to this species on account of the similarity of stereom texture with the superomarginals (Figs. 9b–d, f, h, 10i). The abactinal ossicles of the centre of the disc are flat, with subrounded outlines and the primary ossicles were large, flat and relatively conspicuous (Fig. 10i). The more distal ossicles are smaller, oval, and developed progressively larger internal, laterally directed articular processes. The external faces become smaller distally, and distinctive tall conical ossicles, with actinally directed processes, probably occupied the radial parts of the arms (Fig. 10f). The actinals are flat and rectangular (Fig. 9i), and carry similar rugosities to the inferomarginals.
Adambulacrals are assigned provisionally to this species (Fig. 10h, k). They are block-like and rectangular in actinal aspect, and carry a single large, oval central attachment site for a subambulacral spine. A row of approximately ten small spine bases for the ambulacral spines runs along the adradial margin. The abactinal face carries well-developed articulation structures for the ambulacrals (ada1a,b, ada2, ada3) typical of valvatids (Gale in press). The orals (Fig. 10l, m) have a long, low proximal blade which on the radial face bears bases for oral and suboral spines which are very similar to those on the adambulacrals. The apophyse of the oral has a very large proximal process for the riom muscle. The interradial face of the apophyse has a long, deep notch for the rvg.
Reconstruction and ontogeny:
The large number of marginals available permits reconstruction of the margin and overall form of the species, and some information on the ontogenetic changes in proportions of the marginal ossicles. The arms are only slightly produced, and the interradii slightly concave (Fig. 9a). Small interradial marginals in the material are short and trapezoidal in actinal outline (Fig. 9a, e), with a maximum dimension (length of the inner margin) of 1 mm. They taper towards the lateral margin. With increased size, the marginals become progressively longer and indicate that growth of the species displayed a strong allometry.
Discussion:
The presence of long symmetrical interradial marginals, the elongated, narrow marginals 2 and 3 and the short distal marginals of H. longimarginalis gen et sp. nov. provide a distinctive character combination unique in the family Goniasteridae.
Spinulosida PERRIER, 1884
Pterasteridae PERRIER, 1875
Savignaster GALE, in press
Savignaster wardi GALE, in press
Fig. 12a–i

Savignaster wardi Gale in press. Isolated ossicles, Savigna, all from 2a horizon, except d which is from 1. Oral ossicles in interradial (a) and radial (b) views (BMNH EE 13613, 13614). Abactinal ossicles in abactinal (c, i) and lateral (f) views (BMNH EE 13619, 13620, 13629). d Adambulacral ossicle in actinal view (BMNH EE 13615). g Ambulacral, actinal view (BMNH EE 13618). e Primary interradial, tip of process to show muscle facet (BMNH EE 13632). h Interradial chevron ossicle, abactinal view (BMNH EE 13628). See Gale (in press) for detailed description and illustration. All ×20
2010 Savignaster wardi Gale, pl. 21, figs. 1, 2, 5, pl. 22, figs. 1–3,
pl. 23, figs. 3, 4, 6, 11–12, pl. 24, figs. 1–2, 4, 6, 7–12, 14–16.
Type material:
An individual showing abactinal, adambulacral and ambulacral ossicles from Savigna 2b is holotype (BMNH EE 13610). Paratypes are isolated ossicles (EE 13611-32).
Referred material:
An individual showing spines, and over 300 isolated ossicles, including all major ossicle types.
Description:
The species was figured extensively and described in detail by Gale (in press), and a selection of ossicle types is figured here. The abactinal ossicles which carry a central pedicel (Fig. 12c, f, i), the elongated adambulacrals (Fig. 12d), and the orals (Fig. 12a, b) are particularly distinctive. The only additional ossicle type additional to the suite of material described by Gale (in press) is a first adambulacral ossicle.
Discussion:
Savignaster wardi was identified as a basal pterasterid by Gale (in press), with a combination of plesiomorphic characters found in the paraphyletic stem group “Korethrasteridae”, and some synapomorphies of the Pterasteridae (pedicels on abactinal ossicles, inter-abactinal muscles, highly specialised chevron ossicles and pir).
Family Plumasteridae nov.
Diagnosis:
Multiarmed (12–22 arms) asteroids with broad adambulacrals which occupy the entire actinal surface of the arm and V distally: adambulacrals concavo-convex, 5–8 specialised interlocking articulation ridges and grooves articulate with ridges on adjacent adambulacral (modified ada2-3); abactinal ossicles with numerous lateral projections and embayments, and each carries a central large convex boss with which long, glassy, ridged spines articulate.
Type genus:
Plumaster Wright 1863, is the only genus included. It ranges from the Pliensbachian to the Oxfordian.
Discussion:
The Plumasteridae is established for the distinctive multiarmed genus Plumaster. This is distinguished from other multiarmed spinulosans such as solasterids by the unusual boss-like spine articulations of the abactinal ossicles, and the highly modified adambulacral ossicles, which articulate by means of ridges and grooves.
Plumaster WRIGHT, 1863.
Type species:Plumaster ophiuroides Wright, 1863, OD.
Diagnosis: as for family.
Discussion:
Since the original description of P. ophiuroides (Wright 1863, p. 112, pl. 5, fig. 1), the genus has remained obscure and poorly described. Spencer and Wright (1966) based their description on Wright’s original figures, and placed the genus in the Tropidasteridae. The original specimen of P. ophiuroides from the Pliensbachian of Robin Hood’s or Skinnigrove Bay, Yorkshire, is present in the collection of the Sedgwick Museum, Cambridge (CAMSM J 13784), and is refigured here (Fig. 13). The specimen, preserved in clay, and heavily coated with glue, displays a well-preserved actinal surface revealing adambulacrals, ambulacrals and orals. Ossicles of Plumaster are in fact widespread in Hettangian to Oxfordian fine-grained sediments in the UK, France and Switzerland, but have been assigned to various different asteroids. Thus, Hess (1972) referred a plumasterid adambulacral (op. cit. pl. 2, fig. 1) and an abactinal (op cit. pl. 3, fig. 7) to Terminaster cancriformis, and Villier et al. (2004a, b) figured two abactinal ossicles (op. cit. fig. 3, 8; fig. 4, 13) and an ambulacral (op. cit. fig. 4, 12) as Plesiastropecten halloviensis. Inspection of the type of P. ophiuroides and a new assemblage containing abundant Plumaster material from the Pliensbachian of Sedan, Ardennes, France confirm the correct placement of this material. This material will be described in detail at a later date. The plumasterids are among the morphologically most unusual neoasteroids, and the adambs are convergent with the lateral arm plates of certain ophiuroids (e.g. ophiacanthids) in aspects of their morphology (interlocking ridges between successive adambulacral ossicles, concavo-convex form).

Plumaster ophiuroides Wright, 1863. Holotype, Lower Jurassic, Pliensbachian, Robin Hood’s or Skinningrove Bay, Yorkshire, UK (CAMSM J 13784); a1 actinal surface, a2 enlargement of four radii and part of mouth frame, actinal surface. Note broad, short adambulacrals with transverse single row of large spine bases. Scale bar 1 cm
Plumaster shows some similarities with the five rayed Hettangian genus Plesiastropecten Peyer 1945 in both having broad, short adambulacrals with a transverse single row of spine bases, and the stellate abactinals, each of which carries a single centrally placed boss with which a single spine articulates. However, Plesiastropecten has five rays, whereas Plumaster is multirayed (12–22 arms). Detailed comparison must await description of new material of Plumasteridae.
Plumaster sp. nov.

Plumaster sp. nov. a–e SEM images of ossicles, stenocycloides Subzone, Late Oxfordian, 2b horizon, Savigna. f Early Oxfordian, Q. mariae Zone, Rouges Terres, Switzerland. a Abactinal ossicle; note boss and platform-like areole for articulation and muscle insertion of spine (BMNH EE 13978). b, c Ambulacral ossicles in actinal aspect (EE 13950-1). d Proximal adambulacral in abactinal aspect (EE 13952). e Distal adambulacral in actinal aspect (EE 13953). f Proximal adambulacral in actinal view (EE 13954). a ×35, b–f ×15

Plumaster sp. nov. Drawings of adambulacral ossicles. a Abactinal view, ossicle from Savigna 2b (BMNH EE 13955). b Actinal view of ossicle from Lower Oxfordian of Rouges Terres, Switzerland (EE 13954; see also Fig. 14f). c Reconstruction and interpretation of abactinal face of ossicle, based partially on material from the Pliensbachian davoei Zone of Sedan, Ardennes, France (see Thuy et al., this volume). Scale bar in mm
Referred material:
The Savigna material includes four adambulacrals, five ambulacrals and six abactinal ossicles (BMNH EE 13950-53, 13955; EE 13978). A single ossicle from the mariae Zone of Terres Rouges, Switzerland is also figured (BMNH EE 13954).
Description:
The adambulacrals are thin, concavo-convex, and rectangular–square in actinal outline (Figs. 14d–f, 15a–c). The proximal part of the actinal face carries a transverse row of three to five large spherical spine bass, which extend onto the ad pr (Figs. 14f, 15b). Adjacent adambulacrals imbricated strongly proximally and successive ossicles of a row articulate by means of (4–8) interlocking short ridges and grooves, a highly modified ada2-3. Padam and dadam are short and very broad. The ambulacrals (Fig. 14b, c) are flattened, with heads which carry an elongated, triangular proximal wing. The actinal ridge on the base has a large, rounded articulation with the adamb, and a thin broad flange for dadam and padam. Dentition is lost, and the abtam forms a long narrow strip along the adradial margin. The abactinal ossicles (Fig. 14a) are flattened, stellate, with six to eight processes, and carry a single centrally placed boss-like rounded spine base surrounded by a circular areole for muscle insertion. This resembles the platform of echinoid tubercles.
Forcipulatida PERRIER, 1884
Terminasteridae GALE, in press
Terminaster cancriformis (QUENSTEDT, 1876)
Fig. 16a–f

Terminaster cancriformis (Quenstedt). SEM images of external surfaces of ossicles, Oxfordian, transversarium Zone, Savigna 2a horizon. Terminal in actinal (a1) and abactinal (a2) views (BMNH EE 13636). b Inferomarginal (EE 13737), c radial (EE 13640), and f superomarginal ossicles (EE 13643). Oral in radial (d1) and interradial (d2) aspects (EE 13635). Pir in abactinal view (e). See Gale (in press) for detailed descriptions. a ×20, others ×25
Referred material:
Over 200 isolated ossicles, and a single individual (BMNH EE 13633; EE 13635-46).
Description:
The material from Savigna was described and illustrated by Gale (in press), and a selection of ossicles is figured here. The overall form of the species is well known from essentially complete individuals figured by Hess (1972) and Villier et al. (2009). The arm is constructed of nine ossicle rows including a single radial row, two adradial rows, two superomarginal rows and two inferomarginal rows (Fig. 18a).
Discussion:
This small (R < 25 mm) distinctive species is widespread in Callovian–Tithonian sediments in northwest Europe, and three undescribed Terminaster species are now known from Cretaceous (Hauterivian–Cenomanian). Terminaster represents the stem group to zoroasterids, and possibly to all Forcipulatida (Villier et al. 2009).
Asteriidae GRAY, 1840
Savignasterias gen. nov.
Diagnosis: Asteriidae in which the arm is made up of nine ossicle rows (1 radial, 2 adradial, 2 superomarginal, 2 inferomarginal). Quadriradiate marginals and abactinals are distinctively constructed of coarse anastomosing trabeculae, and a single large sub-central spine base is present on all but the adradial ossicle rows. The sculpture distinguishes Savignasterias from all other asteriids.
Type species: S. villieri sp nov. Oxfordian of the French Jura.
Savignasterias villieri sp nov.

Savignasterias villieri gen et sp. nov. SEM images of external surfaces of ossicles, Oxfordian, transversarium Zone, Savigna, 1 and 2a horizons. Radial ossicles is a holotype (BMNH EE 13956), f paratype (EE 13957). b Adradial (paratype, EE 13958). c Superomarginal (paratype, EE 13959). d Inferomarginal (paratype, EE 13960). e Primary interradial from interradius c–d, fused with two distal abactinals, madreporite missing, represented by hole (EE 13979). ×40

Diagnosis:
As for genus.
Derivation of name:
In honour of the contributions of Loïc Villier (Marseille) to the study of fossil asteroids.
Types:
The radial ossicle figured (Fig. 17a; BMNH EE 13956) is holotype, the other figured ossicles are paratypes (Fig. 17b–f). BMNH EE 13957-60; EE 13979.
Referred material:
18 marginals, radials, adradials, primary interradial, possible adambulacrals.
Description:
The abactinal surface of the arm consisted of nine rows of ossicles (as in Cretasterias and other Mesozoic asteriids—Gale and Villier in press), which formed a reticulate network (Fig. 18). The external surfaces of marginals, adradials and radials all display an distinctive construction of coarse anastomosing trabeculae (resembling a matted root system) immediately distal to the single flat-topped spine base. The radials are more or less symmetrically cruciform, and the proximal process, which imbricates over the tip of the proximal ossicle, is longer than the other three (Fig. 17a, f). A large, flat-topped spine base is positioned proximal to the centre of the ossicle. The adradials are asymmetrically cruciform and lack a spine base (Fig. 17b). The superomarginals are strongly asymmetrical (Fig. 17c), and only the distal process is overlapped by the adjacent sm. The im possess a tall process articulating with the sm, proximal and distal processes, and a short actinal articulation (Fig. 17d). A single pir in the material has similar trabecular construction (Fig. 17e). This is typically asteriid, in that the paired ossicles adjacent to the madreporite are fused with the pir (Gale in press), and the position of the madreporite is marked by an oval hole.
Discussion:
S. villieri compares closely with Cretasterias reticulatus Gale & Villier in press (and other Mesozoic forcipulatids) in the simple reticulate arm construction in which the marginals and abactinals include only nine ossicle rows (Fig. 18c). It differs from C. reticulatus and all other asteriids by the presence of coarse anastomosing trabeculae on the external surfaces of the marginals and abactinals, and the shape of the IM ossicles. Extant asteriids investigated all possess very fine, even trabecular construction on the external face of marginals and abactinals.
Faunal composition and palaeoecology
The Savigna asteroid fauna is unusual in that it is completely dominated by small taxa which achieve adult dimensions with R between 10 and 30 mm; only two taxa (Pentasteria (P.) longispina and Tylasteria jurensis) grew to large sizes (R = 100–150 mm). The fact that taxa known from other localities and different ages (e.g. Savignaster, Jurapecten, Terminaster) are consistently of similar small size can be taken as evidence that this is a more general aspect of Jurassic asteroid faunas from shelfal muddy environments. It is interesting that these taxa belong to, or are closely related with, families which at the present day are characteristically found in the deep sea. For example, benthopectinids only occur below 400 m water depth (except in the Arctic) and pterasterids are typically found in depths exceeding 200 m at low and mid-latitudes (Clark and Downey 1992). Finally, Terminaster is sister taxon to the zoroasterids which only occur in seas deeper than 200 m. Deep sea Mesozoic asteroids faunas are currently virtually unknown, so it is difficult to test the competing hypotheses that (1) present day deep sea asteroid families migrated from the shelves in the late Mesozoic or early Cenozoic, or (2) they were present in both Mesozoic deep sea and shallow environments, but were subsequently displaced from the shelves, probably during the early Cenozoic. A bathyal fauna of Callovian age (depth less than 200 m) includes Terminaster cancriformis (Villier et al. 2009) and thus provides some support for the latter hypothesis.
The trophic assignations of fossil asteroids are difficult to identify, because of the wide diversity of feeding methods employed by some extant families, such as the goniasterids (Jangoux 1982). However, the astropectinid P. (P.) longispina resembles living forms such as Astropecten polyacanthus (Müller & Trotschel) and was also likely to have been an intraoral predator on bivalves, and possibly echinoderms (Jangoux 1982). Most extant asteriids are extraoral feeders on bivalves, and Savignasterias villieri gen et sp. nov. probably had a similar mode of life. Finally, all living goniopectinids and ctenodiscids are deposit feeders on clay grade sediments (Shick et al. 1981; Jangoux 1982; Gale 2005) and it is likely that the goniopectinid Chrispaulia jurassica sp. nov. lived in the same way. This is supported by the presence of an unusually large opening for the first tubefeet on the oral ossicles of Chrispaulia (pers obs); in the living Ctenodiscus crispatus (Retzius), the proximal tubefeet are enlarged and used to shovel sediment into the peristome (Shick et al. 1981).
The surface sculpture of external ossicle surfaces in Chrispaulia jurassica, Jurapecten hessi, Terminaster cancriformis and Savignasterias villieri is made up of coarse trabeculae modified in various ways. On the superomarginals of C. jurassica, irregularly conjoined transverse rows of bead like stereom are present. On the inferomarginals of this species, and all external ossicles of J. hessi and T. cancriformis, small rounded rugosities are joined by fine, radially arranged trabeculae. This distinctive sculpture is similar to that found on arbacioid echinoids, where it is called epistroma. Extant members of the families entirely lack this sculpture (Gale in press). The external faces of ossicles of the asteriid S. villieri are made up of coarse, anastamosing, root-like trabeculae which are not found on any extant members of the family. These surface textures possibly had a defensive function, making the external surfaces of the ossicles resistant to abrasive damage (Table 2).
References
Blake, D. B. (1973). Ossicle morphology of some recent asteroids and description of some West American fossil asteroids. University of California Publications in Geological Sciences, 104, 59 pp, 18 pls.
Blake, D. B. (1986). Some new post-Palaeozoic sea stars (Asteroidea: Echinodermata) and comments on taxon endurance. Journal of Paleontology, 60, 1103–1119.
Clark, A. M., & Downey, M. E. (1992). Starfishes of the Atlantic (794 pp, 118 pls). London: Chapman & Hall.
Desmoulins, C. (1832). Catalogue descriptif des stéllerides vivantes et fossiles. Actes de la Société Linnéenne de Bordeaux, 4, 1–24, 2 pls.
Enay, R. (1966). L’Oxfordien dans la moitié sud du Jura Francais. Nouvelles Archives du Muséum d’Histoire Naturelle de Lyon fasc, 8, 1, 310 pp.
Fisher, W. K. (1911). Asteroidea of the North Pacific and adjacent waters. 1. Phanerozonia and Spinulosa. Bulletin of the United States National Museum, 76, xiii+420 pp, 122 pls.
Forbes, E. (1841). A history of British starfishes and other animals of the class Echinodermata (267 pp). London: Jon Van Voorst.
Forbes, E. (1848). On the Asteridae found fossil in British Strata. Memoirs of the Geological Survey of Great Britain, 2, 457–482.
Gale, A. S. (1987). Goniasteridae (Asteroidea, Echinodermata) from the Late Cretaceous of north-west Europe. 1. Introduction. The Genera Metopaster and Recurvaster. Mesozoic Research, 1, 1–69, 9 pls.
Gale, A. S, (2005). Chrispaulia, a new genus of mud star (Asteroidea, Goniopectinidae) from the Cretaceous of England. Geological Journal, 40, 383–397.
Gale, A. S. (in press). The phylogeny of the Neoasteroidea (Asteroidea, Echinodermata). Special Papers in Palaeontology.
Gale, A. S., & Villier, L. (in press). Mass mortality of an asteriid starfish (Asteroidea: Echinodermata: Forcipulatida): Cretasterias reticulatus n. gen & n. sp. from the Late Maastrictian (Cretaceous) of Morocco. Palaeontology.
Goldfuss, G. A. (1831). Petrefacta Germaniae. Part 3. Dusseldorf.
Gray, J. E. (1840). A synopsis of the genera and species of the Class Hypostoma (Asterias Linnaeus). Annals and Magazine of Natural History, 6(175–184), 275–290.
Heddle, D. (1967). Versatility of movement and the origin of the asteroids. Symposium of the Zoological Society of London, 20, 125–141.
Hess, H. (1955). Die fossilen Astropectiniden (Asteroidea). Schweizerische Paläontologische Abhandlung, 71, 113 pp, 4 pls.
Hess, H. (1966). Mikropaläontologische Untersuchungen an Ophiuren, V: Die Ophiuren aus dem Argovien (unteres Ober-Oxford) vom Guldenthal (Kt Solothurn) und von Savigna (Dept. Jura). Eclogae Geologicae Helvetiae, 52, 2.
Hess, H. (1968). Ein neuer Seesterne (Pentasteria longispina sp. nov) aus den Effingerschichten des Weissensteins (Kt. Solothun). Eclogae Geologicae Helvetiae, 61, 607–614.
Hess, H. (1972). Eine Echinodermen-Fauna aus dem mittleren Dogger des Aargauer Jura. Schweizerische Paläontologische Abhandlungen, 92, 87 pp, 23 pls.
Hess, H. (1999). Seesternplatte vom Weissenstein. Seesterngrabung Schinznach (AG). Sonderdruck aus Schwiezer Strahler, 11(12), 1–15.
Jangoux, M. (1982). Digestive systems: Asteroidea. In M. Jangoux & J. M. Lawrence (Eds.), Echinoderm nutrition (pp. 235–279). Rotterdam: A.A. Balkema.
Perrier, E. (1875). Revision de la collection de Stéllerides du Muséum d’Histoire Naturelle de Paris, 384 pp.
Perrier, E. (1884). Mémoire sur les étoiles de mer recueillis dans la Mer des Antilles et la Golfe de Mexique. Nouvelles Archives du Museum D’ Histoire Naturelle de Paris, 6(2), 127–276.
Peyer, B. P. (1945). Beitrage zur Kenntnis von Rhat und Lias. Eclogae Geologicae Helvetiae, 36, 303–326, 8 pls.
Quenstedt, F. A. (1876). Petrefactenkunde Deuschlands, 1.4, Die Asteriden und Encriniden.
Sahagian, D., Pinous, O., Olferiev, A., & Zakharov, V. (1996). Eustatic curve for the Middle Jurassic and Cretaceous based on Russian platform and Siberian stratigraphy: zonal resolution. Bulletin of the American Association of Petroleum Geologists, 80, 1433–1458.
Shick, J. M., Edwards, K. C., & Dearborne, J. H. (1981). Physiological ecology of the deposit feeding sea star Ctenodiscus crispatus: ciliated surfaces and animal sediment interactions. Marine Ecology Progress Series, 5, 165–184.
Spencer, W. K., & Wright, C. W. (1966). Asterozoans, pp. U4-107. In R. C. Moore (Ed.). Treatise on invertebrate paleontology. Echinodermata. 3 (1). University of Kansas Press: Geological Society of America.
Turner, R. L., & Dearborne, J. H. (1972). Skeletal morphology of the Mud Star, Ctenodiscus crispatus (Echinodermata:Asteroidea). Journal of Morphology, 138, 239–262.
Valette, Dom A. (1929a). Note sur quelques stéllerides Jurassiques. Travaux du Laboratoire de Géologie de la Faculté des Sciences de Lyon, 16, mem 13, 7–30, pls 1–4.
Valette, Dom A. (1929b). Note sur une nouvelle espece d’Astropecten du Bathonien du Tremblay (Nievre). Bulletin de la Société géologique de France 4th série, 29, 415–418.
Védrine, S., & Strasser, A. (2009). High-frequency palaeoenvironmental changes on a shallow carbonate platform during a marine transgression (Late Oxfordian, Swiss Jura Mountains). Swiss Journal of Geoscience, 102, 1–24.
Verrill, A. E. (1899). Revision of certain genera and species of starfishes with descriptions of new forms. Transactions of the Connecticut Academy, 10, 145–234.
Villier, L., Blake, D. B., Jagt, J. W. M., & Kutscher, M. (2004a). A preliminary phylogeny of the Pterasteridae (Echinodermata: Asteroidea) and the first fossil record: Late Cretaceous of Belgium and Germany. Paläontologische Zeitschrift, 78, 281–299.
Villier, L., Kutscher, M., & Mah, C. L. (2004b). Systematics and palaeoecology of middle Toarcian Asteroidea from the “Seuil du Poitou”, western France. Geobios, 37, 807–825.
Villier, L., Charbonnier, S., & Riou, B. (2009). Sea Stars from Middle Jurassic Lagerstatte of La Voulte-sur-Rhone (Ardèche, France). Journal of Paleontology, 83, 389–398.
Ward, D. J. (1981). A simple machine for bulk processing of clays and silts. Tertiary Research, 3, 121–124.
Wright, T. (1863–1880). A monograph of the British fossil Echinodermata of the Oolitic Formations, Vol. II: The Asteroidea and Ophiuroidea. Palaeontographical Society, 1863–1880.
Acknowledgments
My best thanks are to Hans Hess, who introduced me to the Savigna locality, and sent me material he had picked from residues. I would also like to thank David Ward for processing the clay samples, and for help with making plates. Chris Mah kindly advised on extant goniasterids. Dr Simon Cragg and Christine Hughes (University of Portsmouth) assisted with the SEM imagery.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Gale, A.S. Asteroidea (Echinodermata) from the Oxfordian (Late Jurassic) of Savigna, Départment du Jura, France. Swiss J Palaeontol 130, 69–89 (2011). https://doi.org/10.1007/s13358-010-0008-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-010-0008-x