
Abstract
Encrinus liliiformis is a well known crinoid from the middle Triassic Muschelkalk of Europe. A statistical analysis of crown shape was conducted in order to quantify possible differences in crown morphology of E.liliiformis originating from three localities in Southwest Germany (Upper Muschelkalk). These differences basically refer to the length of the brachia and ornamentation patterns on the aboral surfaces of the brachials. The populations originate from different environments: Crailsheim, reflecting a shallow water habitat on a carbonate ramp; Neckarwestheim and Mistlau, originating from a deeper basinal environment. The analysis led to a distinction of two ecophenotypes adapted to the varying habitats: (1) an ecophenotype originating in shallow water showing shorter brachia with less ornamented surfaces of brachials; this type is represented by the Crailsheim population; (2) an ecophenotype originating in a deeper water environment showing longer brachia and a higher degree of ornamentation on brachial surfaces, as represented by the Neckarwestheim population. The crinoids from the Mistlau locality take up an intermediate position in morphology, but the low number of specimens does not allow for statistically significant comparisons. Interpretations of these differences in crown morphology are made with respect to reactions to predatory pressure as well as to variations in water flow energy which is of prime importance for obtaining food resources for these filter feeding animals.
Similar content being viewed by others


Introduction
Phenotypic plasticity is an important and thus intensively discussed subject in functional morphology from both ecological and genetic standpoints. Even for Recent organisms, where detailed information on morphology, living conditions as well as genetics is available, it is still difficult to determine and disseminate the interactions of morphology and environment and potential reasons for differences in phenotypes (for further details see West-Eberhard 2003 or DeWitt and Scheiner 2004). As far as organisms from the fossil record are concerned, only morphological features and palaeoecological data are available to interpret coherences of skeletal morphology and the ambient environment. Genetic information is unknown for the majority of fossils, being restricted to relatively young material preserved under very special conditions. It is thus even more difficult to decide if differences in the morphology of fossil organisms reflect separate species or variations within the reaction norm of the same species showing adaptions to specific ambient environments.
Finding an adequate term for the description of variations in morphology of fossil organisms is not an easy task because of the use of variously defined terms in biology. In this study the term “ecophenotype” will be used to describe the analysed variations in crown morphology of Encrinus liliiformis Lamarck, 1801. The term “ecophenotype” is easily confused with other concepts such as, i.e. “alternative phenotypes” or the general used term “morphotypes” (West-Eberhard 2003). According to Chauffe and Nichols (1995) the term “ecophenotype” should only be used in the context of “… non-persistent, inconsistent, non-inheritable, environmentally induced phenotypic modifications. This would include modifications from disease, injury, physical restrictions on growth or to modifications that develop through use …”. In their study on conodonts, Chauffe and Nichols (1995) use the term “envirotypes” defined as “…persistent, environmentally induced, potentially inheritable phenotypes that have not been genetically selected for an environment”. This does not seem appropriate for the use in this study because the term envirotype is “…a description of factors that are exogenous to the organism. The environmental code and the genetic code together affect the phenotype” (Beckers et al. 2009), but does not describe the phenotype itself. In the present study “ecophenotype” denotes a phenotype (visible physical characteristics or behaviours) that is a result of environmental or ecological conditions rather than genetic expression (nurture instead of nature) (http://Biology-online.org).
Most of the fundamentals of phenotypic plasticity used in functional morphology also apply for ecophenotypic variations. The investigation of ecophenotypes is important for delimiting the morphological range of species variations. Their investigation can also be important for the reconstruction of environmental parameters which control the development and distribution of organisms. These “environmental cues” which induce changes in morphology and behaviour, can include, besides many other parameters, nutrient supply, temperature, light exposure, current regime with flow direction and energy or predation pressure.
Morphological variations of organisms as responses to differences in environment are known from various groups of organisms. The degree of reaction to changes in habitat, which denotes the developmental flexibility (Johnson 1981) is based on the ability of altering body shape during lifetime. Another main factor is the developmental delay, meaning how fast an organism can react to a changing environment (Berrigan and Scheiner 2004). Echinoderms can modify their mesodermal, internal skeleton throughout their entire life. The modular build up of the skeleton, consisting of various distinct plates as well as ongoing addition and resorption of material on the plates leads to an extensive ability of the echinoderms to reflect the ambient habitats by ecophenotypic adaptions.
Environmental heterogeneity, which refers to the time period in which the environment itself changes, is also an important factor. It can be divided into two classes, spatial and temporal heterogeneity (Berrigan and Scheiner 2004). For fossil material, information on the temporal component of environmental heterogeneity in most cases covers several hundreds of thousands to millions of years; a higher resolution of changes is scarce. Spatial variations can be reconstructed if a wide area of deposition can be tracked and facies can be followed laterally.
Crinoids are known since the early Palaeozoic and were widespread in shallow water habitats till the end of the Permian. During the Triassic, the modern Articulata evolved and radiated, including the non-stalked comatulids, which are common inhabitants of Recent shallow water environments. Stalked crinoids, however, shifted in habitat from former shallow water environments to today’s deep water settings during the mid-Mesozoic. This is interpreted in the context of the so called “Mesozoic Marine Revolution” when durophagous (shell-crushing) predators like teleosts, stomatopods, or decapod crustaceans radiated (Vermeij 1977; Meyer and Macurda 1977; Harper 2006). Evidence on the radiation of crinoids following the end-Permian extinction event in connection to benthic predation is also given by Baumiller et al. (2010). Stanley (2008) gives an elaborate summary of the minor role that interspecific competition plays in benthic marine population dynamics compared to the greater influence of predation.
Variations in crown morphologies of E. liliiformis have been known since the nineteenth century and were already described by Beyrich (1858) and Quenstedt (1876), who noticed differences in the occurrence and intensity of knob to spine like swellings of the outer brachial surfaces. Hagdorn (1986) mentions strongly sculptured E. liliiformis specimens as well as crowns which show less sculpturing, but with elongated arms. Encrinus crowns depicted in Hagdorn (2004) display different ornamentation patterns on the secundibrachials. Besides this, the total arm length seems to be increased in specimens that lived in deeper water habitats. An additional difference in morphology is the inflation of radials, primibrachials and the first secundibrachial, which also seems to be greater in specimens originating from deeper water environments. This inflation of distinct plates is also known from Encrinus aculeatus v. Meyer, 1851, originating from the Lower Muschelkalk beds.
In the present study, three populations of E. liliiformis from the Southwest German Upper Muschelkalk beds, from Crailsheim, from Mistlau and from Neckarwestheim, were chosen to determine and quantify these differences in crown shape of the specimens.
Differences in arm morphology and arm length of Recent crinoids are described, e.g. for Chladocrinus (=Neocrinus) decorus and Endoxocrinus parrae (Messing and Llewellyn 1991; Oji 1996). These variations have been interpreted as adaptations to fine-scaled differences in topography, flow regime and predation pressure. Variations in the inflation of calyx plates also appear in Palaeozoic crinoids (Signor and Brett 1984; Gahn and Kammer 2002). Signor and Brett (1984) interpret this tuberosity and the increase in spine formation of Palaeozoic crinoids as a defensive adaptation to predators. David and Roux (2000) found different bathymetric ecophenotypes of Endoxocrinus and Diplocrinus from the tropical western Atlantic and Pacific Ocean, which show differences basically in the design of cirri. Gahn and Kammer (2002) analysed Baryocrinus from the Burlington Limestones (Lower Mississippian), and interpreted the differences in tumidity as primitive (flat) and derived (inflated) conditions.
Materials and methods
Geological setting
During the middle Triassic (244–231 Ma) the supercontinent Pangaea was surrounded by the Panthalassic Ocean and indented by the Tethys. In central Europe, complex tectonic settings caused regional subsidence and a subdivision into several structural highs and grabens (Feist-Burkhardt et al. 2008). During the Anisian, the terrestrial Buntsandstein ultimately changed into the marine conditions of the Muschelkalk (Hagdorn 1991), which lasted from late Bithynian to Fassanian times (Hagdorn and Nitsch 2009).
In the south, the Tethys Ocean was separated from the Central European Basin by the Vindelician–Bohemian Massif (Fig. 1). This led to the different developments of the Germanic and the Alpine Triassic. The Rhenish Massif and the London–Brabant Massif in the west and Fennoskandia in the northeast formed additional areas of non-deposition. Sea level changes and faunal exchange proceeded through the East Carpathian, the Silesian–Moravian, and the Burgundy Gates (Hagdorn 1991; Feist-Burkhardt et al. 2008).

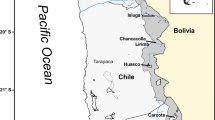
Palaeogeographic reconstruction of the Central European Basin during the Upper Muschelkalk following Hagdorn (1999) with the position of localities: 1 Neckarwestheim, 2 Crailsheim, 3 Mistlau
The Muschelkalk Group is divided in three subgroups: the Lower, the Middle and the Upper Muschelkalk. The Lower Muschelkalk reflects the changing conditions from terrestrial to marine settings. During this time, dolomites and thinly bedded limestones with intercalated shelly and oolitic beds were the main deposits in the western part of the basin, whereas the eastern part is characterized by open marine conditions. At the beginning of the Illyrian, a regression led to the deposition of the mostly evaporitic sediments of the Middle Muschelkalk. In the deepest part of the basin, thick halite deposits are found, laterally interfingering with sulphate- and dolomite-rich sediments, which formed towards the margins of the Central European Basin. The Upper Muschelkalk beds show fully marine conditions caused by a renewed transgression in late Illyrian times. This section consists of thick greyish limestones deposited both in the deeper parts of the basin as well as in shallower coastal regions (marls and fine clastic sediments) (Hagdorn 1991; Feist-Burkhardt et al. 2008; Hagdorn and Nitsch 2009).
The analysed material originates from the Trochitenkalk Formation (moT according to Hagdorn and Simon 2005 and Hagdorn and Nitsch 2009), which was deposited in south western Germany during late Illyrian times (Hagdorn and Nitsch 2009) in a shallow marine setting and is characterized by thick fossiliferous limestones. Facies composition and denotation of beds differs locally, resulting in a number of regional members. Near Crailsheim, the Crailsheim Member consists of massive crinoidal limestones. In more basinal settings of the Hassmersheim Member, individual crinoidal beds, the “Trochitenbänke”, can be distinguished, interbedded with marlstone layers (Hagdorn and Nitsch 2009). In the general stratigraphy of Baden-Württemberg, the Trochitenkalk Formation starts with the first limestone bed above the dolomitic marls of the Middle Muschelkalk. The “Tetractinella-Bank” which contains the brachiopod Tetractinella trigonella is an important ecostratigraphic marker bed. According to the occurrence of the characteristic columnal ossicles of E. liliiformis (“Trochiten”) the “Trochitenbänke 1–11” are additional regional markers. The formation ends with another major marker bed of the Upper Muschelkalk, the “Spiriferina-Bank”, which is dominated by the brachiopod Punctospirella fragilis (Hagdorn and Nitsch 2009).
Crailsheim is situated in the northeast of Baden-Württemberg in Southwest Germany. According to Vollrath (1957), Aigner (1985) and Aigner and Bachmann (1992) the Upper Muschelkalk sediments found in the area around Crailsheim reflect shallow marine conditions on a gently inclined carbonate ramp. Near Crailsheim, E. liliiformis and terquemiid bivalves formed small bioherms above the storm wave base and even above the fairweather wave base with a diameter of 2–3 m and a thickness of up to 1.8 m (Hagdorn 1978). The Crailsheim Member reaches a thickness of up to 16 m (Hagdorn and Nitsch 2009) and is regionally divided into the “Untere Encrinus-Bänke”, “Encrinus-Platten” and “Obere Encrinus-Bänke” (Hagdorn 1978; Hagdorn and Ockert 1993). The analysed Encrinus crowns are derived from the “Encrinus-Platten” and the “Obere Encrinus-Bänke”, which are correlated with “Trochitenbänke 3–6” in the general Muschelkalk marker bed stratigraphy in Baden-Württemberg (atavus- and pulcher-biozones). The four 100 ka-cycles of this interval represent a relatively large time span, such that the “population” of Crailsheim in reality encompasses many generations of distinct populations.
Mistlau, only about 7 km NW of Crailsheim, shows sediments deposited in a deeper environment with a higher content of typical clay and marl beds representing the transition from the shallow water Crailsheim Member to the more basinal Hassmersheim Member (Hagdorn and Ockert 1993). The Encrinus crowns from Mistlau originate from the “Encrinus-Platten”, correlated with Mergelschiefer 2 of the Hassmersheim Member.
Neckarwestheim is located near Heilbronn, circa 70 km to the west of Crailsheim. Around the area of Neckarwestheim, E. liliiformis settled on an area of the sea floor within a deeper part of the Central European Basin (Hagdorn 1999). Here, Encrinus formed loose populations attaching to different bivalve shells and carbonate concretions (Linck 1954). According to Hagdorn (1999) the Encrinus-populations of Neckarwestheim were situated in a position between wave base and storm wave base. Linck (1954, 1965) assumes a water depth of no more than 180 m, based on the presence of symmetric wave ripples. Most of the investigated specimens are found on two slabs exhibited in the “Erdgeschichtliche Sammlung der Firma Märker in Lauffen a. N.” (Fig. 2). The Encrinus bearing layer is “Trochitenbank 6” laterally correlated with the top of the Crailsheim Member (pulcher-biozone). A typical feature of the Neckarwestheim Encrinus specimens is the absence of the upper part of the arms, which may be a consequence of autotomy due to the stress during burial or by predatory action. The Encrinus specimens from Neckarwestheim represent a single population and thus their morphology reflects the living conditions of a short time interval.

Slabs showing part of the Neckarwestheim population, exhibited in the “Erdgeschichtliche Sammlung der Firma Märker in Lauffen a. N.”; rapid burial of the crinoids led to the excellent and rather complete preservation of the organisms; around the area of Neckarwestheim Encrinus liliiformis settled in loose populations in a deeper ramp environment. a Slab SML1001 with six crowns of Encrinus liliiformis, one specimen almost complete with a total length of 1 m; b slab SML1002 with 21 crowns of Encrinus liliiformis, stalk fragments indicate presence of more than 50 individuals; c detail of a showing the typical absence of arm tips either caused by stress-induced autotomy during rapid burial or by predatory action before burial
Determination of crown attributes
Morphological features were determined for all appropriately prepared Encrinus crowns in various collections including: Institut für Geowissenschaften, Universität Tübingen (IGPT), Naturhistorisches Museum Heilbronn (NHMH), Staatliches Museum für Naturkunde Stuttgart (SMNS), Erdgeschichtliche Sammlung der Firma Märker in Lauffen a. N. (SML), Muschelkalkmuseum Ingelfingen (MHI) and the Bayerische Staatssammlung für Paläontologie und Geologie München (BSP). For the analysis, only adult individuals were measured, as recognized by the biseriality of brachia. A total of 56 crowns from Crailsheim, 32 from Neckarwestheim and 11 from Mistlau were thus investigated.
A digital calliper with a resolution of 0.01 mm was used to measure the dimensions of the following crown attributes (Fig. 3): crown length (including radials and all brachials), brachia length (from primibrachial one to the tip of the arm), calyx diameter (outer diameter of radial circlet), and the width and height of radial (R), primibrachial one (IBr1), primibrachial two (IBr2) and secundibrachial 1 (IIBr1) of one radius. The number of brachialia was counted (brachial quantity), differentiating uniserial and biserial IIBrr. Ornamentation intensity (ornamentation intensity) and inflation of RR and IBrr (inflation intensity) could not be measured in a satisfying way. Intensity of ornamentation and inflation were thus classified by dividing each of them into four classes (Fig. 4 showing examples of the four classes of ornamentation intensity): (1) no, (2) slight, (3) clear, and (4) high. The beginning of the ornamentation (ornamentation onset) was determined by counting the first IIBr, where a clear pattern was recognizable. All attributes with respect to brachials were determined for the longest and most complete arm of each of the specimens.

Plate diagram and measured dimensions of Encrinus crowns: R radial, IBr1 primibrachial one, IBr2 primibrachial two, IIBr1 secundibrachial one, arrows indicate location of measurement of width (horizontal arrows) and height (vertical arrows) of the distinct plates

Encrinus liliiformis crowns showing the four different classes of ornamentation intensity: a (1) no ornamentation (MHI1004, Crailsheim population); b (2) slight ornamentation (MHI1213/2, Crailsheim population); c (3) clear ornamentation (GPIT/3/106/168, Crailsheim population); d (4) high ornamentation (SMNS 67684, Mistlau population, note the knob-like ornamentation on the outer arm surfaces)
Statistics
For statistical analysis, the program packages Microsoft Excel 2007 and SPSS 17 were used. Applied statistical methods are: (1) descriptive statistics to analyse composition of populations with regard to the different crown attributes: explorative methods with estimation of arithmetic mean, variance, skewness and kurtosis, frequency with histograms, Gaussian distribution with Shapiro–Wilk test; (2) univariate correlation with comparison of means using the t test for unrelated data to compare the three populations and decide whether the analysed crowns belong to one population or if a distinction is probable; and (3) bivariate correlation with Pearson’s correlation coefficient and Spearman’s rank coefficient to determine variation of crown attributes within the distinct populations.
Results
Descriptive statistics
The data were first tested for normality using the Shapiro–Wilk test. The results determined the appropriate statistical methods to be used. Normality tests of distribution for all measured attributes show significance values of p > 0.05 following the Shapiro–Wilk test except the width of IIBr1 of the Mistlau population. Thus the data points of Crailsheim as well as those of Mistlau and Neckarwestheim follow a Gaussian distribution. The graphs of the Q–Q plots show a linear relationship between the observed values and the expected values from a normal distribution without a clear deviation. All populations are thus treated as normal distributions.
The histograms in Fig. 5 show graphs of normal distribution for crown length of the Crailsheim (left), the Mistlau (centre) and the Neckarwestheim population (right). The skewness of the normal distribution of crown length shows a slight shift to the left. Incorporating juvenile specimens into the statistics for both populations would increase this trend. The kurtosis for the Crailsheim crown length distribution is leptokurtic (kurtosis = −0.429), whereas the populations of Mistlau and Neckarwestheim show a platykurtic distribution (kurtosis = 0.953, respectively 0.151). In general, the Crailsheim and the Neckarwestheim populations show a similar range of crown length with values of 75.78 mm, respectively 78.70 mm. Mistlau has a lower range with a value of 57.80 mm. The smallest of the Crailsheim individuals has a crown length of 34.09 mm, the largest one of 109.87 mm, with a mean of 63.97 mm. Mistlau specimens range from 32.41 mm for crown length to 90.21 mm with a mean value of 62.10 mm. The Neckarwestheim population ranges from 48.24 to 126.94 mm, with a mean of 80.65 mm. All data values of descriptive statistics for the crown length of analysed Encrinus specimens are shown in Table 1.

Histograms of normal distributions shown for crown length; all attributes of the three populations are treated as normal distributions in the following statistic analyses
Univariate correlation
The comparison of means using the t test for unrelated data shows a clear difference between the Crailsheim and the Neckarwestheim population in almost all measured and counted attributes (Table 2). Only the values for height of IBr1, IIBr1 and R indicate an equality of means for both populations. The height of IBr2 with a value of p = 0.045 lies very near to the limiting value of 0.05 which determines the equality/inequality of means within a 95% confidence interval. The population from Mistlau is in some attributes very similar to the population of Crailsheim, in others more similar to that of Neckarwestheim. Only the mean values for ornamentation onset and intensity of ornamentation between the specimens from Crailsheim and Mistlau differ clearly from each other with p = 0.007 and p = 0.013. The comparison of the Mistlau population with that from Neckarwestheim shows significant differences in mean values of crown length (p = 0.004), brachia length (p = 0.005), brachial quantity (p = 0.023) and inflation intensity (p = 0.001).
Bivariate correlation
To test the different attributes for correlation and thus analyse the variation within the populations, bivariate correlation was determined using the Pearson’s correlation coefficient for the measured data and the Spearman’s rank coefficient for the classified data. Results are shown in Table 3. In general, the Neckarwestheim population shows lower values of correlation than the Crailsheim and Mistlau populations.
The width and height of IBr1, IBr2, IIBr1 and R in all populations are correlated within a 99% confidence interval except the width of IBr1 and IIBr1 in the Mistlau population (correlation only within a 95% interval). The width of the distinct plates generally shows higher correlation in all populations than the height, which is a more variable attribute.
Figure 6 shows the corresponding graphic correlation of brachia length plotted against ornamentation onset (left) and against brachial quantity (right). Brachia length and ornamentation onset for the Crailsheim (with R = 0.533) and Neckarwestheim (R = 0.539) populations show a correlation within the 99% confidence interval, Mistlau (R = 0.646) within the 95% interval. The trend lines within the graph display the generally higher values of ornamentation onset in the Crailsheim population and the greater increase in the position where ornamentation starts with increasing arm length. The comparison of crown length–brachial quantity-ratio shows almost equal linear equations for the three populations, the correlation coefficient is very high with R ≥ 0.9. The trend line for Neckarwestheim (dotted) shows the greater arm length and higher number of brachials compared to Crailsheim and Mistlau.

Graphs showing correlation of brachia length and ornamentation onset (left) with higher values for the Crailsheim population compared to Mistlau and Neckarwestheim; and brachia length plotted against brachial quantity (right) displaying a clear and similar linear relationship for all three populations
The calyx diameter and the respective width of the radial is highly correlated with the width of the first IBr (R > 0.9), which directly attaches to the radial. With increasing distance from the radial, the values decrease slightly, but remain high. The calyx diameter is also correlated with crown length. The trend lines display a higher increase in calyx diameter with longer arms in the Crailsheim population compared to that of Mistlau and Neckarwestheim.
The results of bivariate correlation using the Spearman’s rank coefficient for the classified attributes of ornamentation and inflation intensity show generally low values. Only some values of the Crailsheim population are significantly correlated (cf. Table 3). One of these values is the inflation intensity which is positively correlated with the ornamentation intensity in the Crailsheim population. Ornamentation onset and inflation intensity are negatively correlated in the Encrinus crowns originating from Crailsheim.
Discussion
The measurements and brachia countings of Encrinus liliiformis crowns show slight variations between the populations of Crailsheim, Mistlau and Neckarwestheim. Because of the large difference in the number of analysed crowns from the distinct localities, statistical results should be interpreted carefully. First of all, the low number of Encrinus specimens from the Mistlau population leads to a relatively poor significance of statistical results. Another problem is the preservation of Encrinus crowns from Neckarwestheim, where almost all specimens have lost their arm tips. This causes a decreased number of brachials as well as a lower length of brachia. The results in the fact that only the ratio of brachial number–brachia length can provide information about differences between the populations as far as the arms are concerned.
Despite these shortcomings, trends in the statistical distribution of the three distinct populations can be recognized. In general, the Neckarwestheim specimens have longer crowns (mean crown length = 80.65 mm) compared to the crowns originating from Crailsheim (mean crown length = 63.97 mm) and Mistlau (mean crown length = 62.10 mm), which show a more compact appearance. The absence of crowns smaller than 32 mm refers to the selection of adult specimens, while juvenile individuals were not measured. The absence of Neckarwestheim crowns smaller than 48 mm in contrast to the relative high abundance of Crailsheim crowns in this size range with 13 individuals (Mistlau 1 individual) may be a problem related to the smaller sample size of Neckarwestheim. The interpretation of this distribution as a result of ecological adaption would support the observations of Hagdorn (1986, 2004), where the crowns of Neckarwestheim specimens have greater crown length than those of the Crailsheim population.
The results of univariate analyses using the t test for unrelated data show a clear difference between the populations of Crailsheim and Neckarwestheim, whereas Mistlau differs from Neckarwestheim regarding crown/brachia length, brachial quantity and inflation intensity and from Crailsheim in ornamentation onset and intensity. A systematic interpretation leads to the assumption that all specimens of the three localities belong to E. liliiformis without a further distinction of (sub-) species. Even if the crowns originating from Crailsheim and Neckarwestheim differ clearly, the similarity of both populations to that of Mistlau indicates that all analysed specimens belong to one species. The difference in crown length and ornamentation pattern thus represents an ecophenotypic variation of the same species possibly reflecting adaptations to varying habitats.
Bivariate analysis using the Pearson’s correlation coefficient confirms the results of descriptive and univariate analyses. In general, the Mistlau population has more values that show significant linear correlation only within a 95% confidence interval compared to Crailsheim and Neckarwestheim, where, in most cases, the corresponding values lie within the 99% interval. This is related to the small sample size of the Mistlau population. The comparison of crown length–brachial quantity-ratio shows almost equal linear equations for all three populations, indicating that the higher values of crown length in Neckarwestheim specimens originate in an increase of brachial number rather than an increase in thickness of the already existing brachials. The correlation values and the trend lines of height and width of the distinct plates indicate that brachials and radials are more clearly determined with respect to size changes in the Neckarwestheim specimens than in the Crailsheim ones. This may reflect the generally higher variation in habitat in the Crailsheim region, where the Encrinus specimens originate from a wider stratigraphic interval and thus are derived from several distinct populations in contrast to the Neckarwestheim material that represents a shorter time interval and a corresponding lower number of E. liliiformis populations.
The linear correlation of crown length and calyx diameter is high for all populations. The trend line indicates that with increasing crown length, the diameter of the radial circlet increases less in the Neckarwestheim population compared to that of Crailsheim, which supports the more compact appearance of Crailsheim crowns. The elongation of brachia in Neckarwestheim E. liliiformis specimens, interpreted as an ecological adaption, increases the efficiency in food collection with a filtration fan in a deeper basinal environment, where flow energy and nutrient supply may be lower than in a more coastal region like Crailsheim reflected during the middle Triassic.
Beyrich (1858) as well as Quenstedt (1876) already noticed two different morphologies concerning the tumidity of distinct plates and ornamentation on brachial surfaces in E. liliiformis. Beyrich (1858) proposed to establish a new species for the ornamented crowns; Quenstedt (1876) regarded this form as being a different morphotype of the same species. Both authors compared the southern German material to crowns found in Upper Silesia belonging to Encrinus aculeatus. This species shows tumid R and IBr and almost spine like surfaces on the brachia. In contrast to E. liliiformis, however, the basalia overlap the radialia in E. aculeatus and can be seen in lateral view (Quenstedt 1876). In addition, E. aculeatus has cirri on its column (Bielert and Bielert 2000), which are absent in E. liliiformis. The function of the tumidity or the relation to similar morphology of E. liliiformis was, however, not discussed.
Messing and Llewellyn (1991) observed distributional, morphological and postural differences in populations of the recent stalked isocrinids Chladocrinus (=Neocrinus) decorus and Endoxocrinus parrae. These heterogeneities are suggested to reflect fine-scale variations of topography and flow regime. Messing (1994) observed comasteroid crinoids with shorter arms in shallow water habitats and interpreted this feature as a response to flow energy gradients. Even if stalkless crinoids differ in their feeding behaviour by having the ability of changing their feeding place, morphological reaction to flow energy is likely to be similar. Messing et al. (1999) were able to differentiate four different morphs of the genus Endoxocrinus, which may represent different species and reflect differential selective pressure to adapt to the distinct environments. David and Roux (2000) conducted a detailed biometric study of Endoxocrinus and Diplocrinus from the western Atlantic and the Pacific Ocean and found ecophenotypes probably controlled by water flow characteristics. They divided the examined specimens into six empirical morphotypes: four differing in the design of the cirri, one in arm branching pattern and one in the appearance of arms.
Another reason for an increase in arm length in deeper water habitats might be decreased predation intensity (Oji 1996). Though the probability of an attack is proportional to the length of the arm (Oji and Okamoto 1994), the elongation of arms in deeper waters for the benefit of obtaining more particles out of the current is greater than the risk of being attacked by predators. Shorter arms provide less contact surface for predators and are therefore beneficial in shallower waters with higher predation pressure.
The ornamentation differs in its point of origin. Crailsheim, Mistlau and Neckarwestheim specimens show a steady increase in the position of the first IIBr, where ornamentation starts with increasing crown length, but within the populations the starting point is relatively variable. The onset of ornamentation in the Crailsheim population averages at the 27th IIBr, in the Neckarwestheim and the Mistlau population at the 20th IIBr. The ratio of brachia length and ornamentation onset number of IIBr shows only a very low correlation. Trend lines for Crailsheim and Mistlau increase at a steeper rate than the trend line for Neckarwestheim, indicating a greater variability through life than the Neckarwestheim specimens show. The crowns originating from the deeper water habitat around Mistlau and Neckarwestheim have a higher degree of ornamentation leading to more robust arms that may reflect a defence strategy. Signor and Brett (1984) described the development of defensive structures in different groups of Palaeozoic organisms including stalked crinoids. They found a clear increase in the spinosity beginning in the Silurian, which is coeval with a Mid-Palaeozoic radiation of durophageous predators.
Oji (1996) observed two different phenotypes in Recent Endoxocrinus parrae while analysing predation intensity with respect to water depth. In previous studies, these phenotypes were divided into two distinct species, but Oji (1996) regarded them as belonging to the same species. The phenotype living in shallower water shows a smooth dorsal surface on the arms, the phenotype originating in deeper water is characterized by a serrate dorsal surface of the arms. Oji (1996) confirmed the general assumption that in shallower water, predation pressure is greater than in deeper water habitats by determining the regeneration rate of brachia in populations coming from different depths. Oji (1996) analysed a population originating from 150 m down to 750 m depth. The smooth phenotype was found in the whole depth range whereas the serrate phenotype occurred in depths from 400 to 750 m. The functionality of the two different phenotypes is, however, not discussed. The values for water depth of the previously mentioned study on Recent populations are much greater than that assumed for the German Muschelkalk. The locality of Neckarwestheim is presumed to represent a maximum water depth of 180 m (Linck 1954). This difference in distribution may indicate the shift of stalked crinoids from shallow to deep water environments initiated by the Mesozoic Marine Revolution (Vermeij 1977; Meyer and Macurda 1977; Baumiller et al. 2010). In contrast to the Recent E. parrae, E. liliiformis also shows ornamentation in shallow water habitats. An Encrinus crown depicted in Hagdorn (2004) has a smooth surface of brachia and calyx, but during the present study, only one such specimen without any ornamentation pattern was found in the collections.
Baumiller et al. (1999) observed predator–prey interactions between cidaroids and crinoids in Recent populations off the coast of Florida suggesting ecological and evolutionary implications such as escape strategies of crinoids against slow moving predators. Stalk shedding of isocrinids is one possible strategy among others. Baumiller and Gahn (2004) analysed predator-driven evolution in Palaeozoic crinoids by determining the frequency of arm regeneration and showed an increase in arm regeneration suggesting a higher frequency of non-lethal attacks by predators. Antipredatory morphologies became more common with the diversification of shell-crushing predators (Baumiller and Gahn 2004). In their study of fossil material, Baumiller et al. (2010) show that bite marks similar to that of Recent cidaroid echinoids on stalked crinoids occur on a variety of Mesozoic crinoid remains indicating a long existing predator/prey relationship between cidaroids and stalked crinoids, at least from the Triassic onwards.
The function of the ornamentation on the aboral sides of the IIBrr or the swelling of R and IBrr is still not known. Since predation pressure decreases in deeper water habitats (Oji 1996), it is unlikely to be interpreted as a strengthening of arm structure for protection against increased predation. The ornamentation could, however, reflect an adaption to an unknown or unrecognized specialized predator only present in deeper water. From the Upper Muschelkalk deposits of Crailsheim, cidaroids like Serpianotiaris coaeva and other echinoids are known. In Neckarwestheim, the remains of E. liliiformis lost their arm tips before burial. Besides the explanation of autotomy as a reaction to stress, the missing arm tips could also indicate predation just before burial. Possible predators are placodonts or durophagous fish (Linck 1954). Specific examples from the Muschelkalk are the placodont Placodus gigas, the ichthyosaur Tholodus schmidi or the bony fish Colobodus maximus (Schoch and Wild 1999). Ornamentation is visible in all populations, so that a protection against predators in both habitats is expected. As mentioned above, E. liliiformis crowns without any ornamentation is not commonly observed.
The flexibility of the arms of E. liliiformis depends on the number of uniserial IIBrr, which does not differ significantly in the populations. The change from uni- to biseriality has the greatest frequency at fifth to sixth IIBr, but this feature is highly variable even in one specimen. The ornamentation pattern shows no connection to a possible increased flexibility in shallower or deeper water specimens. Another possible explanation for the development of ornamentation on calyx plates and brachials could be an adaption to lower flow energy in deep water environments. The structures on the arms thus lead to a more effective channelling of water flow through the filtering pinnules. This could be accomplished by avoiding or decreasing the formation of turbulences. In deeper water habitats, nutrient supply is not as concentrated as in shallow water environments. In order to obtain enough particles, an increased effectiveness of filter feeding would represent an evolutionary advantage. As shown by Macurda and Meyer (1974) or Roux (1987), the morphology and feeding posture of Recent crinoids depend on current energy as well as on turbulence formation. In slow currents without much turbulence, the filtration fan is opened to its full extent. In higher flow energies, the crinoid opens its filtration fan at lower angles in order to prevent itself from being torn from the substratum. To support the hypothesis of a relationship between morphological variations of E. liliiformis and flow energy, studies on the mode and details of filter feeding are necessary. Computer models and tests in flow channels could clarify a possible effect of the ornamentation to micro-scaled water flow patterns.
Conclusions
In the present study three populations of the middle Triassic crinoid E. liliiformis were compared with respect to their crown morphology. Different attributes were measured including crown and brachia length, dimensions of R, IBr1, IBr2 and IIBr1. Inflation of RR and IBrr and ornamentation onset and intensity of the aboral surfaces of the IIBrr were classified. The results were analysed using various statistical analyses.
Encrinus liliiformis crowns from three localities of Southwest Germany reflecting different life habitats show ecophenotypic variations concerning their crown morphology. The statistical analysis of investigated crowns shows morphological similarities for most of the measured characters; all crowns are thus all deemed to belong to a single species. The differences refer basically to the length of brachia and the ornamentation of IIBrr. The function of these variations is still not known but possible explanations concern the protection against predators or the effectiveness of filter feeding.
The disparities in crown morphology may be the result of various environmental cues such as variations in nutrient supply, flow energy or predation pressure. In deeper water environments, nutrient supply generally decreases. Longer brachia, forming a greater surface for food collection by filter feeding could represent an advantage. Increased ornamentation may be a reaction to different flow regimes, e.g. by canalizing the water for an increased filter feeding effectiveness. The ornamentation increases the stability of the crinoids’ brachia and may therefore be a defence mechanism for better protection against predators.
Encrinus liliiformis was very abundant in the Central European Basin during the late middle Triassic. Future analyses on further specimens from other localities reflecting both shallow ramp and deeper basinal facies would obviously be useful for further analyses of ecophenotypic variations. Crowns from the region around Schwäbisch Hall, for example, appear to have even higher distinction of inflation of RR, IBrr and the first IIBr. In addition, material from northern Germany could be analysed in order to obtain an overview if these adaptations are a regional occurrence or if it is a typical condition for this species throughout the Muschelkalk of Europe.
References
Aigner, T. (1985). Storm depositional systems. Dynamic stratigraphy in modern and ancient shallow-marine sequences. Lecture Notes in Earth Sciences, 3, 174.
Aigner, T., & Bachmann, G. H. (1992). Sequence-stratigraphic framework of the German Triassic. Sedimentary Geology, 80, 115–135.
Baumiller, T. K., & Gahn, F. (2004). Testing predator-driven evolution with paleozoic crinoid arm regeneration. Science, 305, 1453–1454.
Baumiller, T. K., Mooi, R., & Messing, C. G. (1999). Predator–prey interactions between cidaroids and crinoids and their ecological and evolutionary implications. Geological Society of America, Abstracts with Programs, 31, 172–173.
Baumiller, T. K., Salamon, M. A., Gorzelak, P., Mooi, R., Messing, G. G., & Gahn, F. J. (2010). Post-Paleozoic crinoid radiation in response to benthic predation preceded the Mesozoic marine revolution. Proceedings of the National Academy of Sciences, 107, 5893–5896.
Beckers, J., Wurst, W., & Hrabé de Angelis, M. (2009). Towards better mouse models: Enhanced genotypes, systemic phenotyping and envirotype modeling. Nature Review Genetics, 10, 371–380.
Berrigan, D., & Scheiner, S. M. (2004). Modeling the evolution of phenotypic plasticity. In T. J. DeWitt & S. M. Scheiner (Eds.), Phenotypic plasticity, functional and conceptual approaches (pp. 82–97). New York: Oxford University Press.
Beyrich, H. E. (1858). Über die Crinoiden des Muschelkalkes. Abhandlungen der königlichen preußischen Akademie der Wissenschaften zu Berlin, 1858, 1–49.
Bielert, U., & Bielert, F. (2000). Gemeinsames Auftreten von zwei Arten der Gattung Encrinus im Unteren Muschelkalk am Nordrand des Thüringer Beckens. Beiträge zur Geologie von Thüringen, 7, 137–145.
Chauffe, K. M., & Nichols, P. A. (1995). Differentiating evolution from environmentally induced modifications in Mid-Carboniferous conodonts. Palaeontology, 38, 875–895.
David, J., & Roux, M. (2000). Convergence phénotypique chez les pentacrines Endoxocrinus et Diplocrinus liée aux contraintes environnementales. Comptes Rendus de l’Academie des Sciences Serie III—Sciences de la Vie, 323, 281–286.
DeWitt, T. J., & Scheiner, S. M. (2004). Phenotypic plasticity, functional and conceptual approaches. In T. J. DeWitt & S. M. Scheiner (Eds.), (pp. 274). New York: Oxford University Press.
Feist-Burkhardt, S., Götz, A. E., Szulc, J., Borkhataria, R., Geluk, M., Haas, J., Hornung, J., Jordan, P., Kempf, O., Michalík, J., Nawrocki, J., Reinhardt, L., Ricken, W., Röhling, H.-G., Rüffer, T., Török, Á., & Zühlke, R. (2008). Triassic. In T. McCann (Ed.), The Geology of Central Europe Volume 2: Mesozoic and Cenozoic (pp. 749–822). London: The Geological Society.
Gahn, F. J., & Kammer, T. W. (2002). The cladid crinoid Barycrinus from the Burlington Limestone (Early Osagean) and the phylogenetics of Mississippian Botryocrinids. Journal of Paleontology, 76, 123–133.
Hagdorn, H. (1978). Muschel/Krinoiden-Bioherme im Oberen Muschelkalk (mo1, Anis) von Crailsheim und Schwäbisch Hall (SW-Deutschland). Neues Jahrbuch für Geologie und Paläontologie. Abhandlungen, 156, 31–86.
Hagdorn, H. (1986). Neue Funde von Encrinus greppini DeLoriol 1877 aus dem Oberen Muschelkalk von Südbaden und der Nordschweiz. Paläontologische Zeitschrift, 60, 285–297.
Hagdorn, H. (1991). The Muschelkalk in Germany—An introduction. In H. Hagdorn (Ed.), Muschelkalk—a field guide (pp. 7–21). Korb: Goldschneck-Verlag.
Hagdorn, H. (1999). Triassic Muschelkalk of Central Europe. In H. Hess, et al. (Eds.), Fossil crinoids (pp. 164–176). Cambridge: Cambridge University Press.
Hagdorn, H. (2004). Muschelkalkmuseum Ingelfingen (pp. 88). Hagdorn, H. (Ed.), Heilbronn: Edition Lattner.
Hagdorn, H., & Nitsch, E. (2009). 6th International Triassic Field Workshop—field guide.
Hagdorn, H., & Ockert, W. (1993). Encrinus liliiformis im Trochitenkalk Süddeutschlands. In H. Hagdorn & A. Seilacher (Eds.), Muschelkalk. Schöntaler Symposium 1991 (pp. 245–260). Korb: Goldschneck Verlag.
Hagdorn, H., & Simon, T. (2005). Der Muschelkalk in der Stratigraphischen Tabelle von Deutschland 2002. Newsletters on Stratigraphy, 41, 143–158.
Harper, E. M. (2006). Dissecting post-Palaeozoic arms races. Palaeogeography, Palaeoclimatology, Palaeoecology, 232, 322–343.
Johnson, A. L. A. (1981). Detection of ecophenotypic variation in fossils and its application to a Jurassic scallop. Lethaia, 14, 277–285.
Linck, O. (1954). Die Muschelkalk-Seelilie Encrinus liliiformis—Ergebnisse einer Grabung. Naturwissenschaftliche Monatsschrift, Aus der Heimat, 62, 225–235.
Linck, O. (1965). Stratigraphische, stratinomische und ökologische Betrachtungen zu Encrinus liliiformis Lamarck. Jahreshefte des geologischen Landesamtes in Baden-Württemberg, 7, 123–148.
Macurda, D. B., & Meyer, D. L. (1974). Feeding posture of modern stalked crinoids. Nature, 247, 394–396.
Messing, C. G. (1994). Comatulid crinoids (Echinodermata) of Madang, Papua New Guinea, and environs: Diversity and ecology. In B. David, et al. (Eds.), Echinoderms through time (pp. 237–243). Rotterdam: Balkema.
Messing, C. G., Baumiller, T. K., & David, J. (1999). Morphological differentiation in the living isocrinid Endoxocrinus (Echinodermata: Crinoidea): A response to differences in habitat? Geological Society of America, Abstracts with Programs, 31, 171.
Messing, C. G., & Llewellyn, G. (1991). Variations in posture, morphology and distribution relative to current flow and topography in an assemblage of living stalked crinoids. Geological Society of America, Abstracts with Programs, 23, 343–344.
Meyer, D. L., & Macurda, D. B. (1977). Adaptive radiation of the comatulid crinoids. Paleobiology, 3, 74–82.
Oji, T. (1996). Is predation intensity reduced with increasing depth? Evidence from the west Atlantic stalked crinoid Endoxocrinus parrae (Gervais) and implications for the Mesozoic marine evolution. Paleobiology, 22, 339–351.
Oji, T., & Okamoto, T. (1994). Arm autotomy and arm branching pattern as anti-predatory adaptations in stalked and stalkless crinoids. Paleobiology, 20, 27–39.
Quenstedt, F. A. (1876). Petrefactenkunde Deutschlands—Die Asteriden und Encriniden nebst Cysti- und Blastoideen, Abtheilung 1, Band. 4. Leipzig, 1874–1876, pp. 742.
Roux, M. (1987). Evolutionary ecology and biogeography of recent stalked crinoids as a model for the fossil record. In M. Jangoux & J. M. Lawrence (Eds.), Echinoderm studies 2 (pp. 1–53). Rotterdam: Balkema.
Schoch, R., & Wild, R. (1999). Die Wirbeltiere des Muschelkalks unter besonderer Berücksichtigung Süddeutschlands. In N. Hauschke & V. Wilde (Eds.), Trias, Eine ganz andere Welt: Mitteleuropa im frühen Erdmittelalter (pp. 331–342). München: Verlag Dr. Friedrich Pfeil.
Signor, P. W., & Brett, C. E. (1984). The mid-Paleozoic precursor to the Mesozoic marine revolution. Paleobiology, 10, 229–245.
Stanley, S. M. (2008). Predation defeats competition on the seafloor. Paleobiology, 34, 1–21.
Vermeij, G. J. (1977). The Mesozoic marine revolution: Evidence from snails, predators and grazers. Paleobiology, 3, 245–258.
Vollrath, A. (1957). Zur Entwicklung des Trochitenkalks zwischen Rheintal und Hohenloher Ebene. Jahreshefte des Geologischen Landesamtes Baden-Württemberg, 2, 119–134.
West-Eberhard, M. J. (2003). Developmental plasticity and evolution (pp. 794). West-Eberhard, M. J. (Ed.), New York: Oxford University Press.
Acknowledgments
We sincerely thank Hans Hagdorn for his valuable contributions concerning various aspects of this project as well as commenting on an earlier version of this paper. We also thank Uwe Fabich, Wolfgang Hansch, Martin Nose and Günter Schweigert for providing access to museum collections. Wolfgang Gerber is thanked for the excellent photography and Hans Luginsland for specimen cleaning. We sincerly thank Dolf Seilacher and Umberto Nicosia for their constructive reviews.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dynowski, J.F., Nebelsick, J.H. Ecophenotypic variations of Encrinus liliiformis (Echinodermata: Crinoidea) from the middle Triassic Muschelkalk of Southwest Germany. Swiss J Palaeontol 130, 53–67 (2011). https://doi.org/10.1007/s13358-010-0007-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-010-0007-y