
Abstract
The applicability of enzyme immunoassays (EIA) for aflatoxin M1 (AFM1), ochratoxin A (OTA) and zearalenone (ZEN) to analyse these toxins in donkey milk (Equus asinus) was studied. For AFM1 and OTA analysis, milk could be analysed by EIA without sample pretreatment. For ZEN, heat treatment at 78 °C for 30 min prior EIA analysis was required to avoid false positives. To include detection of phase II metabolites of ZEN, samples were additionally treated with glucuronidase/sulfatase for this EIA. Detection limits were 5 ng/kg (AFM1), 9 ng/kg (OTA) and 600 ng/kg (ZEN). All donkey milk samples were negative for all three toxins. Satisfactory quantitation was achieved for spiked samples. Analysis of some cereal-containing donkey feed components (pellets, oats) by EIA revealed absence of aflatoxin B1 (AFB1, < 3 μg/kg) and OTA (< 4 μg/kg), while ZEN was found in pellets (180 μg/kg) and in oats (7 μg/kg). This is the first one study on multitoxin determination in donkey milk by antibody-based test systems. In general, the results confirm that EIAs are convenient tools for mycotoxin detection in donkey milk. However, false-positive results may occur, possibly due to the high lysozyme content of donkey milk, which may exert inhibitory activity in some competitive EIA systems. Therefore, specific validation of each EIA for this specific matrix is required, and re-analysis after heat treatment of EIA-positive donkey milk is highly recommended.
Similar content being viewed by others

Introduction
The total world population of donkeys (Equus asinus) is about 43 million; the vast majority (98%) lives in Africa, Asia and middle/south America (FAOSTAT 2018). In Europe, agricultural use of donkeys for meat and milk production is of minor economic importance. However, in the Mediterranean and in the Balkan areas, donkey meat, donkey milk (powder) and donkey milk cheese (pule) are produced at a small scale. Recently, research interest in donkey milk has increased in Europe, especially in Italy (Camillo et al. 2018). Donkey milk is claimed to be similar to human breast milk in nutrient composition (Silanikove et al. 2016), although analytical data proved otherwise (Guo et al. 2006; Alichanidis et al. 2016; Aspri et al. 2016). Further, it is an alleged alternative in cases of cows’ milk intolerance (Monti et al. 2012; Polidori et al. 2009; Carrocino et al. 2000), but lysozyme which is abundant in donkeys milk recently was identified as a major allergen (Martini et al. 2018).
Donkey milk distinctly differs in composition from cows’ milk or human milk, with very low fat (0.1–1.8%) and protein (1.5–1.8%) contents but a high lactose (5.8–7.4%) content (Guo et al. 2006; Alichanidis et al. 2016). Like mare’s milk, and in contrast to milk from ruminants, donkey milk has a very high content of an inhibitory enzyme, lysozyme, with maximum reported levels up to 1.4 g/l (Salimei et al. 2004) or even 4 g/l (Brumini et al. 2016).
As a monogastric species, the digestive system of equines strongly differs from that of ruminants. Consequently, data obtained from cows concerning mycotoxin carry over into milk are not per se applicable to donkeys. Donkey feed typically consists simply of grass, hay and minerals. In case of professional donkey breeding farms, and for lactating animals, cereals and cereal-based pellets are added to the diet (Aganga et al. 2000). Cereals are a relevant source for mycotoxin intake and consequently might result in a carry-over into milk. In the European Union, maximum levels for mycotoxins in animal feed have only been set for AFB1, at 5 μg/kg (European Communities 2002). For deoxynivalenol, ZEN, OTA, fumonisins (FB1/FB2) and T2/HT2 toxins, the European Commission has published guidance values in various feeds and for several livestock species (European Commission 2006a). Except for fumonisins (5 mg/kg in feed), none of these recommendations specifically is dedicated to equine feeding stuff, and donkeys are not mentioned in this document at all. As a non-ruminant monogastric species, donkeys may be more sensitive towards adverse effects of mycotoxins, but again, little is known here except a specific sensitivity towards fumonisins, the causal agents for fatal leukoencephalomalacia in donkeys (Rosiles et al. 1998; Gross et al. 2015).
Published studies on carry-over from feed and natural occurrence of mycotoxins in donkey milk are scarce and, to our knowledge, most exclusively deal with aflatoxins (Escrivá et al. 2017). Tozzi et al. (2016) fed a diet containing 1 kg maize per day which was contaminated with aflatoxin B1 (AFB1, 202 μg/kg) and aflatoxin B2 (AFB2, 11 μg/kg) to six Romagnola breed donkeys for 12 days. They found low levels of 32 ng/l of AFM1 and up to 23 ng/l of AFM2 in the milk of all animals, and calculated carry-over rates of 0.2% for AFM1 and 0.31% for AFM2. This is much lower than the 2–6% carry-over rates reported for cows (Britzi et al. 2013). In a study on 90 donkey milk samples from seven farms in Greece and Cyprus, Malissiova et al. (2016) did not detect any AFM1 positives with a commercial EIA test kit. More recently, using a commercial AFM1 EIA test kit from another producer, Malissiova and Manouras (2017) obtained AFM1-positive results (5–26.5 ng/l) for 5 out of 36 donkey milk samples from Greece farms. Considering the results of Tozzi et al. (2016), some feed compound ingested by these animals must have been contaminated with AFB1 at very high levels to achieve such results in milk, but unfortunately, Malissiova and Manouras (2017) did not analyse relevant feed components for AFB1, or provided an explanation for their findings. Using yet another commercial EIA for AFM1, Bilandzic et al. (2014) obtained AFM1-positive results of 2–10 ng/l in donkey milk from Croatia (14 samples), but did not report the number of positives. An AFM1 EIA test kit from a fourth producer was used to analyse milk from donkeys in Serbia, and 3 out of 5 samples were AFM1 positive, at levels of 3–35 ng/kg (Kos et al. 2014). Although neither Bilandzic et al. (2014) nor Kos et al. (2014) included feed analyses in their studies, these two surveys were possibly influenced by high AFB1 contamination of maize in some Balkan countries in 2012 (Kos et al. 2013) which subsequently lead to elevated AFM1 levels in cows’ milk. This is supported by a recent study (Cammilleri et al. 2018) which found no AFM1 in 84 samples of donkey milk from Sicily, Italy, during 2013–2016.
For ZEN, only one study (Capriotti et al. 2015) analysed donkey milk by liquid chromatography-mass spectrometry (UHPLC-ESI-MS/MS), but all five samples were negative at a claimed detection limit of 20 ng/kg.
In this situation, a more specific assessment of the situation with regard to mycotoxins in donkey milk seems to be required. Commercial mycotoxin EIA test kits for mycotoxins in milk are available for only for AFM1. These have been validated for cows’ milk, in some cases for sheep and goats’ milk, but not for equine milk. However, it is questionable if validation data for cows’ milk are applicable to donkey milk because it is a strongly differing matrix. For other mycotoxins, there is a complete absence of EIA application studies using donkey milk. Therefore, the aim of this study was to assess the applicability of three mycotoxin EIAs for AFM1, OTA and ZEN for analysis of donkey milk, as part of a project to establish simple and versatile screening systems for residues and contaminants in equine milk.
Materials and methods
Sample materials
Samples were obtained from a small herd of Baudet du Poitou donkeys in Germany. In this herd, six mares were lactating. Each one milk sample (about 50 ml) of the six lactating donkeys, which had been collected during routine herd management, was available for analysis. The average age of these animals was 7.5 ± 2.7 years; the average body weight was 350 kg. During this study, the feed of the animals consisted of grass and hay ad libitum. Additionally, a single batch of each a commercial pelleted feed (containing oat fibre, wheat bran and maize), purchased from a local distributer and oat grains from a non-commercial source in the area of Giessen, Germany, were fed to these animals. The amount of these feeds consumed per animal per day was not controlled, but was estimated 0.2–0.3 kg of pellets and 0.5 kg oats per day. While grass and hay were not analysed for mycotoxins, a 3-kg sample of each pellets and oats was collected. This sample was manually mixed in a 10-l plastic bucket, and a subsample of 500 g was collected. These samples were ground to a mean particle size of < 1 mm in a laboratory mill, and manually mixed again before each series of analyses. These samples were analysed for OTA and ZEN by competitive direct EIA as described previously (Liesener et al. 2010; Gross et al. 2015), while the EIA for AFB1 (Gathumbi et al. 2001) was used with a sample preparation method described by Märtlbauer et al. (1991). Neither AFB1 (detection limit 3 μg/kg) nor OTA (detection limit 4 μg/kg) was found in pellets and in oats, the ZEN-EIA determined 180 μg/kg in pellets and 7 μg/kg in oats.
EIA analysis
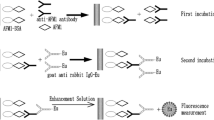
Polyclonal rabbit antisera and toxin-horseradish peroxidase conjugates for competitive direct EIAs for ZEN and OTA were used as described previously (Usleber et al. 1992; Schneider et al. 2001). For AFM1, a commercial enzyme immunoassay test kit (R-Biopharm, Darmstadt, Germany, R1111) was used according to manufacturer’s instructions, except that milk samples were not defatted before analysis. For analysis of AFM1 and OTA, 50 μl of milk sample per well were directly analysed by EIA without any further purification or dilution.
For ZEN-EIA analysis, preliminary studies had indicated the likelihood of false-positive results when using raw milk samples. Therefore, milk samples were heated at 78 °C in a water bath for 30 min, and then cooled to room temperature for analysis of ZEN. Also, exclusively for the analysis of ZEN, a 2 ml portion of milk was mixed with 24 μl of ß-glucuronidase/sulfatase (Sigma Aldrich, G7017, approx. 2200 U/ml milk), incubated for 2 h at 37 °C, and then diluted 1:6 with EIA buffer (5% acetonitrile/PBS pH 7.2). Fifty microlitre per well were then analysed by the ZEN-EIA.
The EIA standard curve cutoff values (detection limit, LOD) for OTA and ZEN were set at a toxin concentration resulting in 20% binding inhibition compared to the negative control (IC20), corresponding to the end of the semilinear range of the standard curve. Four replicates of all standard and sample solutions were analysed, and each milk sample was analysed in at least two different dilutions, to check for sample matrix effects. For the AFM1 EIA, the detection limit was equal to the standard with the lowest toxin concentration (5 ng/kg). For this test, milk samples were analysed in duplicate wells only.
To study sample matrix interferences, milk samples were artificially contaminated with different concentrations of toxin standard solutions before analysis.
Results and discussion
All three EIAs enabled sensitive and simple detection of the respective mycotoxin analysed. With regard to AFM1, the detection limit of 5 ng/kg was five times lower than the lowest maximum level (25 ng/kg) set by European Union regulation 1881/2006 in milk-based food for infants (European Commission 2006b). For OTA, no specific regulation according to European Union regulation 1881/2006 for milk exists, but the lowest maximum level set for “dietary foods for special medical purposes intended specifically for infants” is at 500 ng/kg. The detection limit of the OTA EIA (9 ng/kg) clearly was below that. For ZEN, which is not regarded as a relevant contaminant in cows’ milk (Kappenstein et al. 2005), the lowest maximum level according to European Union regulation 1881/2006 of 20 μg/kg is set for “processed cereal-based foods (excluding processed maize-based foods) and baby foods for infants and young children”. Again, the ZEN-EIA enabled detection of this toxin in donkey milk well below this level, with a detection limit of 600 ng/kg.
All six samples were clearly negative for AFM1 and OTA, which is plausible considering the negative results for AFB1 and OTA in pellets and oats, which were part of the donkeys’ diet, and assuming that grass and hay are not major sources of intake for toxins. For the AFM1-EIA, no further validation was done within this study, but recovery of AFM1 from mares’ milk at levels of 5–15 ng/kg had been found at 91–133% (unpublished results). In the OTA-EIA, donkey milk yielded absorbance values around the standard curve blank (B0), as shown in Fig. 1. Donkey milk spiked at 100, 200 and 400 pg/kg yielded toxin results of 44–77% of the nominal values, with coefficients of variation ranging from 4.3–20%. Currently, no information is available on the risk of OTA contamination in donkey milk, but it has been found in blood of 83% of horses at levels up to 700 pg/ml (Minervini et al. 2013), which suggests that transfer into milk of lactating donkeys is possible depending on the intake of this toxin. The OTA-EIA could be a convenient tool to screen donkey milk samples.

Comparison of EIA absorbance readings for blank and corresponding spiked milk samples from six donkeys in the EIAs for ZEN and for OTA. Spiking level was 2000 ng/kg for ZEN, while for OTA, three different spiking levels (100, 200 and 400 ng/kg) were tested. Not all samples were spiked with each concentration of toxin
For the ZEN-EIA, preliminary experiments had shown that all six-donkey milk samples resulted in binding inhibition of about 30–40% (60–70% B/B0), even for diluted samples. Since such results seemed to lack plausibility in the light of low to moderate ZEN levels in pellets and oats, milk sample matrix interference was considered as a possible cause. Although the ZEN-EIA has successfully been used and validated for the analysis of ZEN in raw cows’ milk (Usleber et al. 1992), the high levels of lysozyme, which is a protein with antimicrobial properties, in donkey milk were assumed to be a possible cause of the observed inhibition. Lysozyme is practically absent in the milk of cows, sheep and goats, but levels up to 4 g/l have been found in donkey milk (Brumini et al. 2016). Lysozyme is quite heat stable, and 62 °C for 30 min or 90 °C for 1 min did not eliminate it from milk (Coppola et al. 2002), temperatures higher than 60 °C may even increase activity due to dissociation of di- and trimers (Vincenzetti et al. 2017). In contrast, Ozturkoglu-Budak (2018) reported a 60% decrease of lysozyme at 85 °C for 2 min. Although no explanation of the possible mode of action of lysozyme on enzyme immunoassay could be provided here, heating donkey milk at 78 °C for 30 min strongly weakened the competitive binding inhibition in the ZEN-EIA and increased absorbance readings to about 80–85% B/B0. Such sample matrix effects seemed to be test specific, since they were not observed in the AFM1 EIA and the OTA EIA. Therefore, it could be due to effects on antibody binding rather than on toxin-horseradish peroxidase activity, but further studies are required on this phenomenon. Dilution of heat-treated samples 1:6 with buffer solution yielded absorbance values close to the standard curve blank value (B0), as shown in Fig. 1. Results did not change with or without addition of glucuronidase/sulfatase, indicating that phase II metabolites of ZEN were not present at relevant levels in these samples. Artificially, contaminated milk samples at a level of 2000 pg/kg were clearly positive (Fig. 1), resulting in ZEN-EIA results of 62 ± 7% of the nominal value. At present, it is unknown whether or not ZEN could be an issue in donkey milk. In comparison with a standard curve detection limit of some improvement of the sensitivity in milk seems to be desirable for this assay. The standard curve detection limit of 100 ng/kg provides sufficient sensitivity to achieve some further improvement.
The observations made on the putative lysozyme–related matrix effects on the ZEN-EIA should be considered for the evaluation of previously reported results for AFM1 in donkey milk. Based on the results of Tozzi et al. (2016) who showed that carry-over of AFB1 as AFM1 into donkey milk is minimal compared with cows’ milk, positive AFM1 results in milk require very high contamination levels of feed compounds which are fed in addition to grass and hay. Results as reported by Malissova and Manouras (2017) or by Kos et al. (2014) require very high AFB1 levels in feed exceeding 100 μg/kg, a level which hardly could be unobserved for prolonged periods, also in the light of European Union regulations (5 μg/kg) for AFB1 in feed for lactating animals (European Communities 2002). Assuming that lysozyme could also negatively affect commercial competitive mycotoxin EIAs in a similar way as the ZEN-EIA, control analyses are advisable for such tests. Reanalysis of positive samples after heat treatment, for example, 78 °C for 30 min as used in this study, would be one option. Further, positive results in donkey milk are certainly not a routine occurrence in food hygiene but demand mycotoxin analysis in feed, to identify possible sources of the toxin. A third option would be the confirmation of positive results by independent methods, which may be more difficult in case of AFM1 than for OTA or ZEN. In any case, EIA validation data obtained for cows’, sheep or goats’ milk are of limited value for donkey milk analysis. Control tests, including analysis of confirmed negative samples and spiked negative samples, and checking for effects of heat treatment, are essential when donkey milk should be analysed for mycotoxins by enzyme immunoassay techniques.
References
Aganga AA, Letso M, Aganga AO (2000) Feeding donkeys. Livestock research for rural development 12, Article #11. http://www.lrrd.org/lrrd12/2/agan122.htm. Accessed 10 Aug 2018
Alichanidis E, Moatsou G, Polychroniadou A (2016) Composition and properties of non-cow milk and products. In: Tsakalidou E, Papadimitriou K (eds) Non-bovine milk and milk products. Academic Press, Amsterdam, pp 81–115
Aspri M, Economou N, Papademas P (2016) Donkey milk: an overview on functionality, technology and future prospects. Food Rev Int 33:316–333
Bilandzic N, Bozic D, Dokic M, Sedak M, Kolanovic BS, Varenina I, Cvetnic Z (2014) Assessment of aflatoxin M1 contamination in the milk of four dairy species in Croatia. Food Control 43:18–21
Britzi M, Friedman S, Miron J, Solomon R, Cuneah O, Shimshoni JA, Soback S, Ashkenazi R, Armer S, Shlosberg A (2013) Carry-over of aflatoxin B1 to aflatoxin M1 in high yielding Israeli cows in mid- and late- lactation. Toxins 5:173–183
Brumini D, Criscione A, Bordonaro S, Vegarud GE, Marletta D (2016) Whey proteins and their antimicrobial properties in donkey milk: a brief review. Dairy Sci Technol 96:1–14
Camillo F, Rota A, Biagini L, Tesi M, Fanelli D, Panzani D (2018) The current situation and trend of donkey industry in Europe. J Equine Vet Sci 65:44–49
Cammilleri G, Graci S, Collura R, Buscemi MD, Vella A, Macaluso A, Giaccone V, Giangrosso G, Cicero A, Lo Dico GM, Pulvirenti A, Cicero N, Ferrantelli V (2018) Aflatoxin M1 in cow, sheep, and donkey milk produced in Sicily, Southern Italy. Mycotoxin Res, in press. https://doi.org/10.1007/s12550-018-0329-y
Capriotti AL, Cavaliere C, Piovesana S, Stampachiacchiere S, Samperi R, Ventura S, Laganà A (2015) Simultaneous determination of naturally occurring estrogens and mycoestrogens in milk by ultrahigh-performance liquid chromatography–tandem mass spectrometry analysis. J Agric Food Chem 63:8940–8946
Carrocino A, Cavataio F, Montalto G, D’Amico D, Alabrese L, Iacono G (2000) Intolerance to hydrolysed cow’s milk proteins in infant: clinical characteristics and dietary treatment. Clin Exp Allergy 30:1597–1603
Coppola R, Salimei E, Succi M, Sorrentino E, Nanni M, Ranieri P, Belli Blanes R, Grazia L (2002) Behaviour of Lactobacillus rhamnosus strains in ass’s milk. Ann Microbiol 52:55–60
Escrivá L, Font G, Manyes L, Berrada H (2017) Studies on the presence of mycotoxins in biological samples: an overview. Toxins 9:251. https://doi.org/10.3390/toxins9080251
European Commission (2006a) Commission recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding (text with EEA relevance) (2006/576/EC). Off J Eur Union L229:7–9
European Commission (2006b) Commission regulation (EC) no 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off J Eur Union L364:5–24
European Communities (2002) Directive 2002/32/EC of the European parliament and of the council of 7 May 2002 on undesirable substances in animal feed. Off J Eur Comm L140:10–27
Food and Agricultural Organization database [FAOSTAT] (2018) search for “world” and “number of asses”. available from:http://faostat3.fao.org. Accessed 2018.08.09
Gathumbi JK, Usleber E, Märtlbauer E (2001) Production of ultrasensitive antibodies against aflatoxin B1. Lett Appl Microbiol 32:349–351
Gross M, Bauer JI, Wegner S, Friedrich K, Machnik S, Usleber E (2015) Mycotoxicoses in horses: toxins, occurrence in feed, and the search for markers of exposure. In: Lindner A (ed) Equine nutrition and training conference (NUTRACO) 2015. Wageningen Academic Publishers, The Netherlands, pp 43–58
Guo HY, Pang K, Zhang XY, Zhao L, Chen SW, Dong ML, Ren FZ (2006) Composition, physicochemical properties, nitrogen fraction distribution, and amino acid profile of donkey milk. J Dairy Sci 90:1635–1643
Kappenstein O, Brockmeyer A, Dietrich R, Curtui V, Klaffke H, Lepschy J, Märtlbauer E, Schneider E, Seidler C, Thielert G, Usleber E, Weber R, Wolff J (2005) Zearalenon in Lebensmitteln. Mycotoxin Res 21:79–82
Kos J, Mastilovic J, Hajnal JE, Saric B (2013) Natural occurrence of aflatoxins in maize harvested in Serbia during 2009-2012. Food Control 34:31–34
Kos J, Lević J, Duragić O, Kokić B, Miladinović I (2014) Occurrence and estimation of aflatoxin M1 exposure in milk in Serbia. Food Control 38:41–46
Liesener K, Curtui V, Dietrich R, Märtlbauer E, Usleber E (2010) Mycotoxins in horse feed. Mycotoxin Res 26:23–30
Malissiova E, Arsenos G, Papademas P, Fletouris D, Manouras A, Aspri M, Nikolopoulou A, Giannopoulou A, Arvanitoyannis IS (2016) Assessment of donkey milk chemical, microbiological and sensory attributes in Greece and Cyprus. Int J Dairy Technol 69:143–146
Malissiova E, Manouras A (2017) Monitoring aflatoxin M levels in donkey milk produced in Greece, intended for human consumption. World Mycotoxin J 10:203–206
Martini M, Swiontek K, Antonicelli L, Garritani MS, Bilò MB, Mistrello G, Amato S, Revets D, Ollert M, Morisset M, Hilger C (2018) Lysozyme, a new allergen in donkey’s milk. Clin Exp Allergy (in press). https://doi.org/10.1111/cea.13232
Märtlbauer E, Dietrich R, Terplan G (1991) Erfahrungen bei der Anwendung von Immunoassays zum Nachweis von Mykotoxinen in Lebensmitteln. Arch Leb 42:3–6
Minervini F, Giannoccaro A, Nicassio M, Panzarini G, Lacalandra GM (2013) First evidence of placental transfer of ochratoxin A in horses. Toxins 5:84–92
Monti G, Viola S, Baro C, Cresi F, Tovo PA, Moro G, Ferrero MP, Conti A, Bertino E (2012) Tolerability of donkey’s milk in 92 highly-problematic cow’s milk allergic children. J Biol Homeost Agents 26:75–82
Ozturkoglu-Budak S (2018) Effect of different treatments on the stability of lysozyme, lactoferrin and blactoglobulin in donkey’s milk. Int J Dairy Technol 71:36–45
Polidori P, Beghelli D, Mariani P, Vincenzetti S (2009) Donkey milk production: state of the art. Ital J Anim Sci 8:677–683
Rosiles MR, Bautista J, Fuentes VO, Ross F (1998) An outbreak of equine leukoncephalomalacia at Oxaca, Mexico, associated with fumonisin B1. Zentralbl Veterinaermed A 45:299–302
Salimei E, Fantuz F, Coppola R, Chiofalo B, Polidori P, Varisco G (2004) Composition and characteristics of ass’s milk. Anim Res 53:67–78
Schneider E, Usleber E, Dietrich R, Märtlbauer E (2001) Entwicklung eines hochempfindlichen Enzymimmuntests zum Nachweis von Ochratoxin A. Mycotoxin Res 17A:170–173
Silanikove N, Leitner G, Merin U (2016) Influence of animal health, breed, and diet on non-cow milk composition. In: Tsakalidou E, Papadimitriou K (eds) Non-bovine milk and milk products. Academic Press, Amsterdam, pp 61–79
Tozzi B, Liponi GB, Meucci V, Casini L, Dall’Asta C, Interore L, Gatta D (2016) Aflatoxins M1 and M2 in the milk of donkeys fed with naturally contaminated diet. Dairy Sci Technol 96:513–523
Usleber E, Renz V, Märtlbauer E, Terplan G (1992) Studies on the application of enzyme immunoassays for the Fusarium mycotoxins deoxynivalenol, 3-acetyldeoxynivalenol, and zearalenone. J Veterinary Med Ser B 39:617–627
Vincenzetti S, Cecchi T, Perinelli DR, Pucciarelli S, Polzonetti V, Bonacucina G, Ariani A, Parrocchia L, Spera DM, Ferretti E, Vallesi P, Polidori P (2017) Effects of freeze-drying and spray-drying on donkey milk volatile compounds and whey proteins stability. LWT Food Sci Technol 88:189–195
Acknowledgements
We thank Margit Kessler, Renate Stumpf and Stephanie Steitz for their excellent technical Assistance. Furthermore, we wish to thank the Brigitte and Wolfram Gedek foundation for financial support of this study.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
None.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Gross, M., Ploetz, C.P. & Gottschalk, C. Immunochemical detection of mycotoxins in donkey milk. Mycotoxin Res 35, 83–87 (2019). https://doi.org/10.1007/s12550-018-0333-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12550-018-0333-2