
Abstract
Lachish (Tell ed-Duweir) is located in the southern part of the Judean foothills, known as the Shephelah, and is one of the larger and most extensively excavated multi-period sites in the southern Levant. We present the faunal results of the first three seasons of the most recent excavations, the Austrian-Israeli Expedition to Tel Lachish. The expedition focusses on two areas of the tell encompassing the Middle Bronze Age III through the Iron Age II, area S (deep section) and area P (palace area). The aims for the faunal analysis are threefold: comparing the results between the two areas, seeing how our results compare to previous analyses, and comparing Lachish to other synchronous sites in the Shephelah. We observe differences in subsistence strategies between the areas in addition to diachronic differences. Ovicaprids dominate all assemblages, but we see shifts in the sheep to goat ratio and mortality profiles through time indicating changes in subsistence strategies. Our new results largely agree with the results from previous analyses, showing the value of previous studies and their potential compatibility with newer research. A synchronic comparison of Lachish within the Shephelah shows the occupants of the site were largely self-sufficient but possibly engaged in an exchange of resources in the vicinity.
Similar content being viewed by others

Introduction
Lachish (Tell ed-Duweir, 31° 33′ 54″ N, 34° 50′ 56″ E) is one of the larger tells in the southern Levant, covering 12 ha, and lies in the southern part of the Judean foothills, referred to as the Shephelah. Scholars believe the site was settled as early as the Pottery Neolithic (Ussishkin 2004), but in this study, we concentrate on the Middle Bronze Age (MBA, 2000–1550 BCE), Late Bronze Age (LBA, 1550–1200 BCE), and Iron Age II (IA II, 975–586 BCE) (period ranges follow Kamlah and Riehl in press). The political importance of Lachish during the Bronze and Iron Age is confirmed by the Assyrian annals describing its conquest by Sennacherib depicted in the Lachish reliefs in his palace in Nineveh (Ussishkin 1982), the El-Amarna letters (Cochavi-Rainey and Rainey 2015), and Papyrus Hermitage 1116A (Golénischeff 1913; Webster et al. 2019).
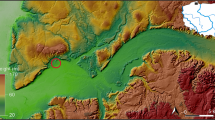
We discuss the faunal remains from the Austrian-Israeli Expedition to Tel Lachish (2017–2019), led by Streit and Höflmayer, in the framework of the ‘Tracing Transformations’ project. The excavation focusses on two areas of the tell area S (deep section) and area P (palace area) (Fig. 1). Area S contains a long stratigraphic sequence from the LBA to the IA representing a settlement area (Table 1): strata S-1 and S-2 contain several pits and densely laminated layers rich in seeds, and stratum S-3 contains a monumental building consisting of several walls. Area S contains settlement assemblages, and Ussishkin (2004) found a public building accompanied by domestic structures in this part of the site. Area P provides a sequence of the MBA and LBA (Table 1): strata P-6 to P-3 contain the MBA palace, whereas strata P-2 and P-1 have domestic architecture dating to the LBA (Streit et al. 2018).

Map of Lachish showing the location of the excavation areas P and S
We have three goals for our analysis of the faunal material. First, we want to establish how subsistence strategies changed through time and whether we can determine differences between areas S and area P during the LBA II. Second, we investigate whether our results establish similar trends as previous analyses. Third, we compare the results of Lachish with other synchronous sites in the Shephelah region to determine how an important centre such as Lachish fits within the regional framework. Answering these questions will allow us to gain a better understanding of subsistence during the Bronze and Iron Age at Lachish and in the broader region.
Background
After the ‘collapse’ of settlements in the Early Bronze Age IV, the MBA I was a period of renewed urbanization in the southern Levant during which Lachish was resettled (Tufnell 1958; Ussishkin 2014). Major architectural finds include the remains of a palace, and structures belonging to a cultic place, although archaeologists have found no images or cult objects (Tufnell 1958; Ussishkin 2004). During the MBA II, Lachish developed into a city-state, and a new palace was built in the centre of the tell. The most noticeable architectural feature is the massive fortifications surrounding the tell (Burke 2008). At the end of the MBA, the palace, and probably the entire city-state, was destroyed by a fire of an unknown cause. Afterwards, the site was home to a secondary settlement, though eventually Lachish was abandoned (Tufnell 1958; Ussishkin 2004).
Overall, the environment was semiarid, but stable oxygen isotope evidence from the speleothems at Soreq Cave indicates considerable fluctuation in palaeo-rainfall patterns throughout the whole Bronze Age which was of a relatively humid character (Bar-Matthews and Kaufman 1988). The δ18O oscillations throughout this stage are characterized by four short dry spells, the last of which falls into MBA I.
During the LBA I Lachish was resettled, although it is not certain to what extent. The Fosse Temple, which existed throughout the LBA, was built, though as with the MBA II, scholars have recovered no images or cult objects. Bietak (2002) suggested the space would have been used as a place of gathering for funerary meals. At this time, the fortifications of the MBA were out of use.
During the LBA II, ancient Egypt became dominant in the region, and, under its patronage, Lachish became an important city-state. Around 1200 BCE, Lachish was again destroyed by fire, though it was rebuilt shortly thereafter (Tufnell 1958; Ussishkin 2004). Although the Fosse Temple was not restored, the Acropolis Temple and the Pillared Building, a large public structure, were constructed. Lachish grew into one of the largest cities in Canaan and prospered. Occupants developed overland and marine trade routes, with the presence of saltwater fish indicating trade with the Mediterranean areas (Lernau and Golani 2004). We can see Egyptian influence in the material culture (Koch 2014; Ussishkin 2004). Around 1130 BCE, Lachish was destroyed and burned by an unconfirmed enemy (Tufnell 1958; Ussishkin 2014).
The LBA pollen record from Galilee documents a similarly high number of Mediterranean tree species as already in MBA II-III with a sharp decline towards the end of the LBA, which may be in relation to the documented drop of the Dead Sea level, and with generally reduced settlement activity in region (Langgut et al. 2015).
After about two centuries, Lachish was resettled in the IA IIA. During this time, Lachish became the second most important city in Judah. This is attested to by the construction of a fortress city with a large civilian population. In 701 BCE, Lachish was destroyed by the Assyrian king Sennacherib (Ussishkin 2004, 1977). After an occupation gap, the site was briefly resettled before it was destroyed for the last time in 588/586 BCE by the Babylonian king Nebuchadnezzar (Tufnell 1953; Ussishkin 2004).
Stable carbon isotope evidence from IA Tel Burna, about 20 km south of Lachish indicate optimal soil moisture availability in the surroundings of the settlement, increasing throughout the IA IIB and IIC (Riehl and Shai 2015).
Although it is currently not possible to link the general environmental patterns to the settlement history of Lachish, it cannot be excluded that observable deteriorating environmental fluctuations may have contributed to the end of some of the settlement phases.
Previous work at Lachish
The first expedition to Lachish was the Wellcome-Marston Expedition (1932–1938), directed by Starkey, whose main goal was to put Lachish in a cultural and chronological framework. The results are published in three volumes by Tufnell (Tufnell 1958, 1953; Tufnell et al. 1940). Bate analyzed the faunal remains (Bate 1958, 1953), and Baden-Powell analyzed the shells (Baden-Powell 1958). Unfortunately, we cannot compare the results of these faunal analyses to our results since the older data lack the resolution of modern faunal analyses.
In 1966 and 1968, Aharoni (1975) excavated at Lachish, focussing on structures dating to the Persian period. Though Lernau (1975) analyzed the fauna of the sanctuary, most material is outside the temporal scope of our study (Late Bronze Age: NISP = 53, Iron Age: NISP = 298).
The longest endeavour at Lachish was the ‘Renewed Excavations’ directed by Ussishkin from 1973 until 1994. The main aim was to investigate the period of the Judean monarchy. The results are published in five volumes (Ussishkin 2004) and a public interest book (Ussishkin 2014). Several zooarchaeologists analyzed the material (Bar-Yosef Mayer 2004; Croft 2004a, 2004b; Drori and Horowitz 1989; Koch 2014; Lernau and Golani 2004). The analyses included fauna from area P and area S, which gives us a baseline for comparing the results from our current study.
From 2013 until 2017, Garfinkel, Hasel, and Klingbeil excavated at Lachish, focussing on the northern and north-eastern edge of the tell and west of the Solar Shrine. They have published some of their results (Garfinkel et al. 2019a, 2019b; Sass et al. 2015; Weissbein et al. 2019, 2016), but the faunal analysis is still in progress.
In 2015 and 2016, Ganor directed two seasons of excavation focusing on the IA gate complex for the Israeli Antiquities Authority. No faunal results have been published yet.
Methods
We analyze faunal remains from hand-collection and flotation from areas P and S recovered during the 2017, 2018, and 2019 seasons of the Austrian-Israeli Expedition. We present the data for the MBA III, LBA II, and IA II (merging IA II A, B, and C to increase the sample size). We measure specimens using Von den Driesch (1976) with 0.0-cm precision and weigh with 0.01-g precision using a digital scale. We identify specimens to skeletal element and bone portion, and assign them to the genus and species level when possible, otherwise to body size class (e.g. large mammal) (Stiner 2005). We focus on macro-mammalian remains, since these contributed most to animal husbandry practices. We choose NISP (number of identified specimens) as our primary quantification unit but calculate MNI (minimum number of individuals) and MNE (minimum number of elements) (Lyman 2018, 2008). We assess diversity in our assemblage by using the reciprocal of Simpson’s index, a measure of taxonomic evenness (Simpson 1949). When calculating 1/D, we only use species-specific identifications, with the exception of sheep and goats, which we combine into an ovicaprid category. To determine differences in taxonomic composition between samples, we perform a Pearson chi-square test using taxa determined to species.
We record bone surface modifications (Behrensmeyer 1978; Fisher 1995; Lyman 1994; Stiner et al. 1995) and evaluate density-mediated attrition to check for preservation bias by comparing the MNE of the most frequent lower tooth with mandibular MNE (Stiner 1991). We also check for recovery bias, since most of the material was hand-collected. We do this by looking at histograms representing the bone length in millimetres for the most common taxon, in this case ovicaprids, per sample. In our histograms, we use bins representing a 5-mm range. We examine body part profiles by dividing the body into nine anatomical regions: horn, head (cranium and mandible), neck (atlas, axis, cervical vertebrae), axial column (thoracic, lumbar vertebrae, sacrum, ribs, and innominate), upper front limbs (scapula and humerus), lower front limbs (radius and ulna), upper hind limbs (femur), lower hind limbs (tibia, astragalus, and calcaneum), and feet (metacarpal, metatarsal, phalanges) (Stiner 1991). Within these regions, we calculate minimal animal units (MAU) by dividing the observed number of elements by their expected numbers in a complete skeleton. This standardizes our MNE counts so we can examine anatomical profiles of major taxa. In our analysis, we assigned axial elements such as ribs; cervical vertebrae 3-5; and thoracic, lumbar, and caudal vertebrae to body size class (e.g. medium ungulate, large ungulate). In calculating our anatomical profiles, we provide species-specific profiles and also profiles based on body size class to make sure that elements underrepresented for a given species were not simply assigned to a general size class category.
We perform ageing using epiphyseal fusion, tooth wear, and tooth eruption. We use epiphyseal fusion of all long bones to calculate survivorship profiles for ovicaprids (Moran and O’Connor 1994) and cattle (Habermehl 1975). We represent survivorship in percentages using a non-cumulative line graph. We also analyze age at death using tripolar graphs, where we plot the frequency of juvenile, prime age adult, and old adult ungulates (Greenfield and Arnold 2008; Steele and Weaver 2002; Stiner 1990; Weaver et al. 2011). Juvenile animals are defined by deciduous fourth premolars. Prime age and old adults are differentiated by wear on fourth premolars, with old adults having more than half of the tooth surface worn off.
We differentiate sheep and goat morphologically (Boessneck 1969; Zeder and Lapham 2010; Zeder and Pilaar 2010) and morphometrically (Davis 2017) when possible in order to establish the sheep to goat ratio, which helps us interpret herding strategies. Keeping a mixed flock of both sheep and goats provides herd security, a common strategy to reduce risk since sheep and goats have different requirements and susceptibilities. For example, when disease strikes only half of the herd will be affected (Redding 1981; Sasson 2016). In addition to this, goats are flexible feeders that browse and graze, have lower water requirements, and reproduce faster than sheep (Redding 1981, 1984). Sheep are less versatile than goats in the sense that they are grazers, but they can provide wool (Redding 1981, 1984). Therefore, fluctuations in the sheep to goat ratio along with changes in their mortality profiles might indicate climatic constraints, or might reflect a preference for meat, milk, or wool production (Payne 1973; Redding 1981, 1984). Socio-political organization also may influence the sheep to goat ratio (Perevolotsky et al. 1989; Redding 1984). In addition to calculating the sheep to goat ratio, we determine sex ratios in ovicaprids using mixture analysis, which uses biometric data to determine the bimodality that most likely reflects sexual dimorphism (Monchot et al. 2005). In ovicaprids, males are typically larger than females due sexual size dimorphism, though sexual dimorphism is more pronounced in goats than in sheep (Davis 2000).
We could not differentiate pig from boar due to the small sample sizes and a lack of suitable skeletal elements at Lachish.
Results
In zooarchaeological assemblages, there is a well-known relationship with larger sample sizes and higher richness (i.e. number of species) (Lyman 2008), which can make it difficult to compare between contexts with different sample sizes. We checked the effect of sample size on richness in our assemblages at Lachish, which shows the relationship is not significant (Pearson’s correlation, r = 0.410, p > 0.05). Since the r value is high enough, this could indicate a mild relationship so we performed individual rarefaction to check our result (SI 2.1). This indicates sample size and diversity are independent, so differences we see in the faunas are not because of variation in sample size (see SI 1 for a complete list of taxa per area and subperiod).
Species abundance
Area S
Area S is mostly represented by the LBA II (Table 2). The assemblage is dominated by ovicaprids, with goats outnumbering sheep (sheep to goat ratio = 0.34, see also SI 2.2 for morphometric data). The low number of wild taxa shows hunting played a minor role. The small presence of suids indicates occasional consumption. Dogs are overrepresented by NISP, due to the presence of an almost completely recovered young dog, reflected in the MNI value. The assemblage has a low diversity (1/D = 1.85) because of the large number of ovicaprids.
Area P
Area P is mostly represented by MBA III, LBA II, and IA II (Table 3). During the MBA III, the most common remains are ovicaprids, with a near-equal presence of sheep and goat (sheep to goat ratio = 0.88). Fish were an important part of the diet, followed by cattle. Suids were consumed moderately. Wild taxa are present in small numbers, as are birds and shells. The diversity is low (1/D = 1.64), reflecting the dominance of ovicaprids.
Ovicaprids are the most abundant group in the LBA II assemblage, with sheep outnumbering goats (sheep to goat ratio = 1.94). Suids were moderately consumed. There are more wild animals in the MBA III assemblage, and fish have a relatively high abundance. The LBA II has low diversity (1/D = 2.00) and is dominated by ovicaprids. The taxonomic composition between the MBA III and LBA II in area P is significantly different (Pearson’s chi-square = 16.80, DF = 7, P < 0.05, Cramer’s V = 0.19).
In addition to the large number of ovicaprids in the IA II assemblage (sheep to goat ratio = 0.70), cattle formed a large part of the diet. No suids are present in this assemblage, and wild animals only play a small role. The diversity is low because of the dominance of ovicaprids (1/D = 1.2). The assemblages between the IA II and LBA II at area P (Pearson’s chi-square = 35.84, DF = 5, P < 0.05, Cramer’s V = 0.26) are significantly different.
Comparisons between areas S and P
Though the two areas are largely similar, with an abundance of ovicaprids and occasional evidence for suid consumption, there are a few differences between the areas in certain time periods. During the LBA II, areas P and S have a significantly different taxonomic representation from one another (Pearson’s chi-square = 598.39, DF = 10, P < 0.05, Cramer’s V = 0.85).
Body part representation
During the LBA II in area S, ovicaprids are most represented by lower hind limbs, front limbs, and head elements (Fig. 2a). Axial, neck, and feet are present in low numbers. Cattle (Fig. 2b) are mainly represented by front and hind limbs. Axial, neck, and feet elements are rare in the assemblage. Anatomical profiles for ovicaprids combined with medium ungulates and cattle combined with large ungulates can be found in SI 2.3, which confirm the pattern we see here.
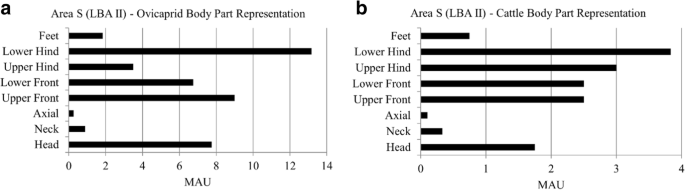
Body part representation for ovicaprids (a) and cattle (b) during the LBA II in area S
MAU values are quite small for area P. During the MBA III in area P, we see feet elements are underrepresented in ovicaprids (Fig. 3a). The pattern continues during the LBA II (Fig. 3b) and during the IA II in area P (Fig. 3c). Anatomical profiles for ovicaprids combined with medium ungulates can be found in SI 2.4, which confirm our patterns. Due to a small sample, we cannot reconstruct anatomical profiles for cattle in area P.
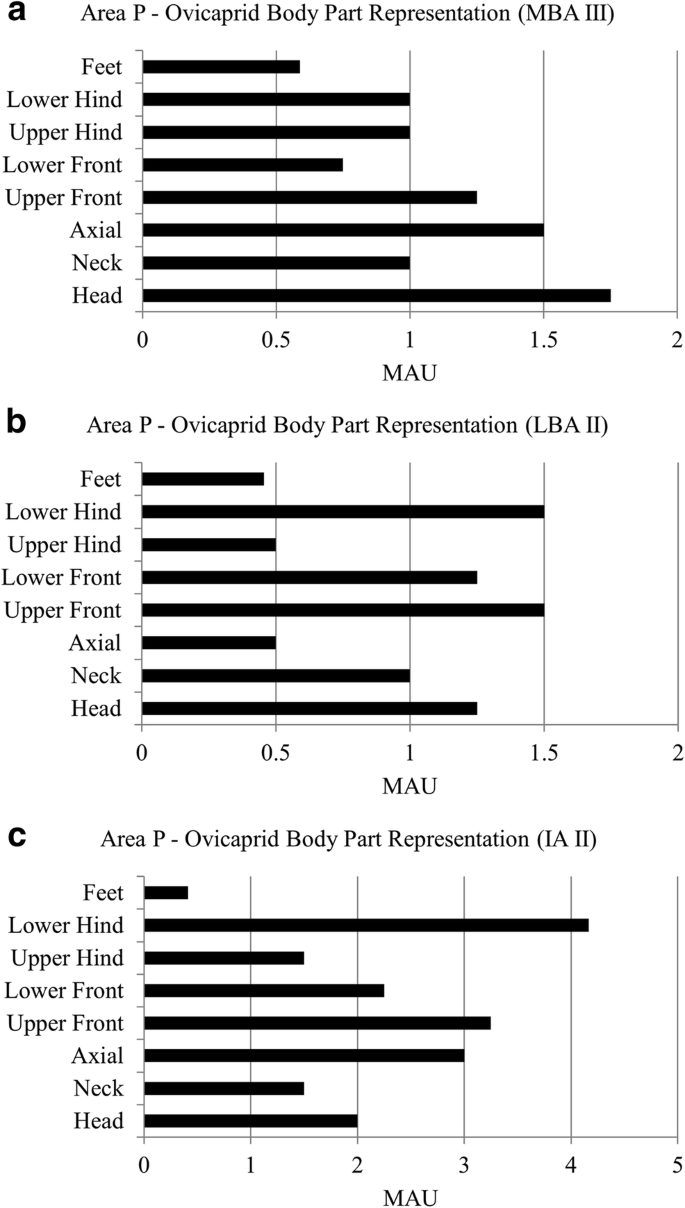
Body part representation for ovicaprids through time in area P. A: body part representation during the MBA III, B: body part representation during the LBA II, C: body part representation during the IA II
Ageing and sexing
We present epiphyseal fusion data for ovicaprids (Fig. 4a, Table 4) for the LBA II in area S. The majority of ovicaprids survive to be sub-adults, with a major drop in survival in adulthood. This corresponds to the results of the tripolar graph, which show the presence of mostly juvenile animals (Fig. 4b). Epiphyseal fusion for cattle (Fig. 5, Table 5) shows most cattle survive the first 2 years of life, with a major drop in survival at year 3, although we should consider the small sample size.
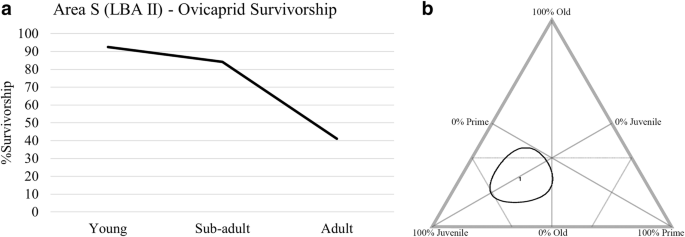
Ageing for ovicaprids during the LBA II in area S. A: %survivorship graph based on epiphyseal fusion. B: tripolar graph represented by 29 juvenile, 14 prime aged adult, and 13 old specimens
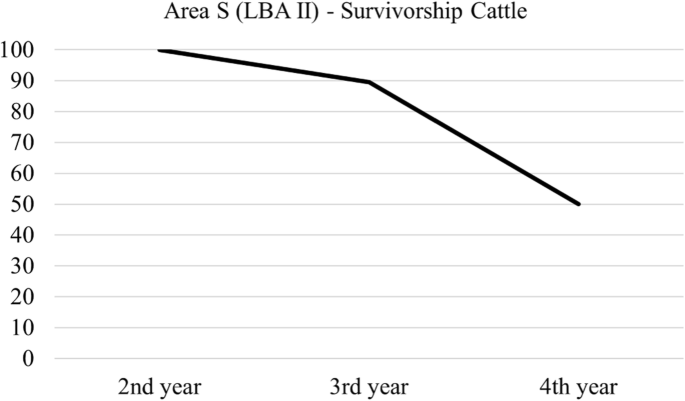
Ageing for cattle during the LBA II in area S
During the MBA III in area P, most ovicaprids (Fig. 6, Table 6) reached sub-adulthood, with a steep drop in survival in adulthood. During the LBA II, the pattern of ovicaprid survival (Fig. 6, Table 6) is similar, but with a higher survivorship in adulthood. The sample size of ageable elements for ovicaprids during the IA II is too small to be evaluated. The same applies to cattle in area P.
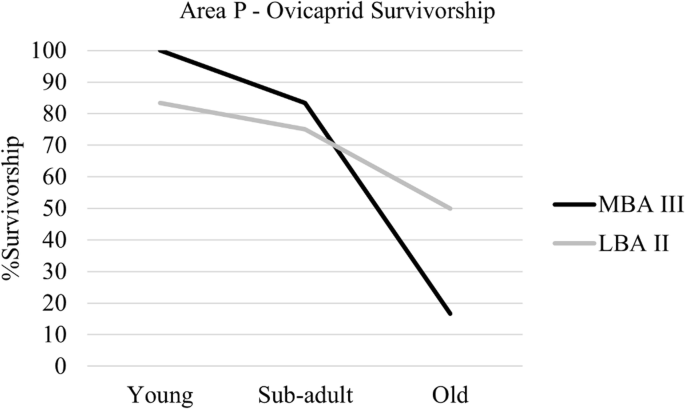
Ageing for ovicaprids in area P through time
We determine the sex of ovicaprids for the LBA II in area S by using measurements of the distal breadth of the first phalanx (Von den Driesch 1976) to perform mixture analysis (N = 27, mean = 11, standard deviation = 2.20; Fig. 7, Table 7). The measurements have a normal distribution (Shapiro-Wilk test, W = 0.943, p = 0.1443). The mixture analysis shows bimodality, with most specimens belonging to the group with the larger mean, indicating males outnumber females (AIC bi-model: 73.18, AIC tri-model: 76.74).

Mixture analysis for ovicaprids during the LBA II in area S
Taphonomy
We checked for the effect of density-mediated attrition in our assemblages (Table 8). We find an even tooth to bone ratio during the LBA II in area P, and an almost even tooth to bone ratio during the IA II in area P and during the LBA II in area S, indicating density-mediated attrition did not affect these assemblages and the trends we see reflect human subsistence strategies. In the MBA III of area P, the tooth to bone ratio indicates a higher preservation of bone compared to teeth. Since most of the material is hand-collected, we also checked analyzed effects of recovery bias by analyzing the bone length of the most common taxon, ovicaprids (SI2.5). It seems recovery bias is not an issue for the LBA II sample in area P and area S, since both these areas have most bone fragments in the smallest sized bins (5–20 mm). This is, however, not the case for the MBA III and IA II in area P, where most fragments are in larger bins (20–40 mm). Recovery methods were the same in the two areas, but the IA II contained the lowest number of materials coming from flotation samples which explains why we do not have that many bone fragments in the lower sized bins. Although the MBA III had more flotation samples than the LBA II in either area, the number of specimens in all MBA III flotation samples was lower than the LBA II samples. Nonetheless, this slight bias should not affect our interpretations too much, since we have flotation samples for all assemblages and bone fragments of 20 mm are still large enough to include most elements from ovicaprids.
Butchery evidence is rare at Lachish (Table 9). Forty-two bones show traces of butchery in area S, most of them dating to the LBA II. All LBA II cutmarks are on ovicaprids, most of them on the humerus and astragalus. Five bones in area P show traces of cutmarks.
Burning is uncommon (1.1% carbonized, 0.3% calcined, Table 10) in area S; most is from the LBA II and occurs on medium ungulates and ovicaprids. Area P has low numbers of burned material (1.0% carbonized, 0.5% calcined), most occurring on MBA III and LBA II material. The MBA III has burning on medium mammals and ovicaprids; the LBA II has most burning on medium mammals and large ungulates
In area S, four LBA II specimens show pathologies and seven bones were worked by humans. Area P has two pieces of worked bone.
Discussion
Subsistence at Lachish during the MBA, LBA, and IA II
During the MBA III, the diet was dominated by ovicaprids, with sheep and goat playing an equally important role. Mixed flocks are more resistant than single taxon flocks, for example against disease or temperature changes (Redding 1981). Sheep and goat were slaughtered as they reached adulthood and their maximum size. Most ovicaprid remains consisted of head elements but the high abundance of lower limbs is noteworthy since these do not contain a lot of meat. Fish and cattle were important parts of the diet. Cattle remains are mostly represented by meaty limb elements. Suids were moderately consumed, and hunting did not play an important role in the diet.
The LBA II is represented by area S and area P. The assemblages in both areas are dominated by ovicaprids; in area S, goats outnumber sheep, whereas in area P, we see the opposite. We should note the small sample size for the ovicaprid differentiation in area P, which probably influences the sheep to goat ratio. Ovicaprids in both areas show similar mortality profiles, with a preference for adult animals. In both areas, ovicaprids are mostly represented by front limbs. Sex ratios of ovicaprids dating to the LBA II in area S show male ovicaprids were slaughtered more than females. Both areas show cattle were an important part of the diet at the site. In area S cattle tended to be slaughtered as adults, indicating their use as traction animals (Bartosiewicz et al. 1997). Suids did not play an important role in the LBA II diet at Lachish, nor did wild animals. Fish played a smaller role in the diet compared to the MBA.
The IA II is dominated by ovicaprids, with goats slightly outnumbering sheep, indicating a strategy focussed on herd security (Redding 1981, 1984). This seems unrelated to environmental developments, which show the IA II occurred during favourable conditions for agriculture.
Comparison to previous analyses
We compare our results for the LBA II in area S to those of Croft (2004a). He divided the LBA into two subperiods, LBA II which is represented by strata in area S, and LBA III which is from two levels represented in several areas of the site. The bulk of the LBA material from Croft (2004a) came from area S, but there is also a larger sample from area P. His assemblage was dominated by ovicaprids, with the sheep to goat ratio remaining stable between subperiods, although there was a slight predominance of goats, like our results for area S. The mortality profiles differ slightly between our study and Croft’s (2004a). Croft (2004a) found that more sheep than goats reached adulthood, since sheep were kept for their wool. Goats tend to be slaughtered at a younger age for obtaining milk (Croft 2004a). Our (smaller) sample is less nuanced and shows most ovicaprids are slaughtered as adults.
He concluded that areas P and S only differ in the ovicaprid to cattle ratio. Our assemblage also shows a difference in ovicaprid to cattle ratio between the two areas, keeping in mind the different sample sizes. Taxa such as pig, birds, and wild animals are represented in low numbers in both assemblages. Fish are present in larger numbers than in the MBA for the previous faunal work, and in similar proportions as our results (Lernau and Golani 2004).
Synchronic comparison of subsistence patterns in the Shephelah
We compare Lachish to other sites in the Shephelah region dating to the MBA, LBA, and IA II (Fig. 8). In doing so, we lose chronological resolution by broadening the time periods to gain more comparative material. We compare sites by looking at the sheep to goat ratio and their mortality profiles (Table 11). The sheep to goat ratios from Lachish in Table 11 differ from the values found in our study due to the larger sample sizes and changes in chronological resolution.

Map showing the location of Lachish and the sites used for comparing subsistence patterns
During the MBA, there is a clear preference for sheep over goats at Lachish. During the MBA, mostly adult animals were slaughtered, with sheep surviving into adulthood more frequently than goats, whereas goats were slaughtered more often at a young age for milk production (Croft 2004a). The smaller sample of our study shows most ovicaprids are slaughtered upon reaching adulthood. At Tell Miqne/Ekron and Tel Haror, sheep outnumber goats. Ovicaprids of all ages are slaughtered at Tell Miqne/Ekron, indicating an unspecialised economy (Maher and Hesse 2016b). At Tel Haror, mostly young animals were slaughtered for obtaining milk and meat products (Klenck 2002). Interestingly, Tell Nagila is the only site in the region to have no sheep but only goat, although Ducos (1968) mentions the possibility of two specimens perhaps being sheep. Mortality data show most goats were slaughtered between 1 and 3 years and only a few were older.
During the LBA, the sheep to goat ratio at Lachish was similar to that of Tel Beth-Shemesh and Timna, with sheep and goat being almost equally important indicating a focus on herd security. At Lachish, we see the same mortality pattern as during the later MBA (Croft 2004a). Ovicaprid mortality at Tel Beth-Shemesh (Tamar et al. 2013) and Timna (Lernau 1988) shows a focus on obtaining meat and secondary products. Tell Miqne/Ekron had sheep outnumbering goats, with a preference of slaughtering young and old ovicaprids. This indicates an unspecialised economy (Lev-Tov 2010, 2000). The occupants of Tel Burna show a strong preference for goats, but we have no age data available to check for economic strategies (Greenfield et al. 2017). The choice for goats could be connected to the arid climatic conditions prevailing during the LBA (Drori and Horowitz 1989; Langgut et al. 2015; Rosen 1986). Greenfield et al. (2017) mention the size of Tel Burna indicates it participated in regional politics. Since Lachish is only about 10 km away, there might have been an exchange of resources between the two sites.
During the IA II, the sheep to goat ratio at Lachish has slightly more sheep than goats and its economic strategy remains the same as during the late MBA and LBA. A focus on sheep husbandry during the IA II fits well with the archaeobotanical evidence in the Shephelah region. At Tel es-Safi a large number of grape pips have been found in IA II contexts (Mahler-Slasky and Kislev 2012), and occupants of Tel Burna cultivated demanding crop species, such as flax, which indicated high water availability (Orendi 2020). This interpretation is further supported by the stable carbon isotope data of barley grain (Riehl and Shai 2015). Tel Halif has more goats, and initially, the focus was on slaughtering prime adults for their meat. In the eighth century, this shifted to slaughtering both prime adults and older individuals for meat and secondary products and eventually slaughtering all age groups (Sapir-Hen 2015). The focus on goats at Tel Halif could be because the site is located close to the northern Negev region, with more arid conditions to which goats are better adapted. On the other hand, this could also reflect that management strategies at Tel Halif were less market-orientated and more for the consumption of people at the site (Sapir-Hen 2015). Tell Miqne/Ekron and Tell el-Hesi had sheep outnumbering goats. Age data at Tell Miqne/Ekron indicates a preference for slaughtering prime adults but keeping enough stock alive into adulthood to profit from their secondary products, such as wool (Lev-Tov 2010, 2000; Maher and Hesse 2016). At Tell el-Hesi, mostly prime adults were slaughtered, indicating an economy focussed on meat (Peck-Janssen 2006).
Conclusion
In this study, we present new data on areas S and P at Lachish. First, we wanted to establish how subsistence strategies changed through time at Lachish and whether we could determine differences between the two areas during the LBA II. Ovicaprids dominate all assemblages, although we see changes in the sheep to goat ratio. During the MBA III in area P, we see an almost equal presence of sheep and goat, indicating a strategy aimed towards herd security. During the LBA II in area S, which is characterized as being a settlement area, we see goats outnumbering sheep, with more male than female ovicaprids being slaughtered. This contrasts with the LBA II area P, which contains palace and settlement remains, where sheep outnumber goats. This could be an artefact of sample size, but perhaps in the future, these differences will help us interpret the ways in which the two areas were used. The IA II has goats slightly outnumbering sheep. We see a preference for slaughtering adult ovicaprids during the MBA III and LBA II. Throughout the history of Lachish, cattle played an important role in subsistence, although we see a sudden decrease in their abundance during the IA II. Fish were an important part of the diet, especially during the MBA III and the LBA II in area P. Suids were moderately consumed during the MBA III and LBA II. Wild animals appeared in low numbers, indicating hunting was not important for subsistence or the economy. The animals were likely hunted locally since they consist of taxa such as fallow deer or gazelle, which are native to the environment around Lachish.
Second, we investigated whether our results establish similar trends as previous analyses. Though we could only compare the LBA II from our results with the LBA results (representing the LBA II and III) from Croft (2004a), our results correspond well with those of the previous analyses. This gives us confidence that previous studies done by different analysts will be useful one day for reconstructing even broader subsistence patterns at Lachish.
Third, we compared the results of Lachish with other synchronous sites in the Shephelah region to determine how the site fits within the regional framework. During the MBA, the ovicaprid economy is similar to other sites in the region, namely sheep outnumbered goats, with an unspecialised economy. During the LBA, subsistence strategies at Lachish fit in the regional framework, being similar to Timna and Tel Beth-Shemesh, with a focus on herd security. This pattern might also reflect drier environments during the end of the LBA (Drori and Horowitz 1989; Langgut et al. 2015; Rosen 1986). The large size of Tel Burna suggests that it might have participated in regional politics which probably involved interactions with Lachish. Tell Miqne/Ekron is the only site in the region where sheep outnumber goats. Most sites during the LBA in the Shephelah tend to focus on a combination of obtaining meat from prime adults and secondary products from older individuals. During the IA II subsistence strategies at Lachish differ from the other sites in the region. Whereas the occupants of Lachish focussed on obtaining secondary products, people at other sites in the region focussed more on meat by slaughtering prime adults.
Lachish was one of the largest tells in the southern Levant, and its political importance, both on a regional and supra-regional level, is attested to by several sources (Cochavi-Rainey and Rainey 2015; Ussishkin 2004). By studying its faunal remains, we can reconstruct subsistence strategies more in-depth and get a better idea of how such a large settlement managed to feed its inhabitants. We demonstrated the merit of synthesizing previous and ongoing faunal analyses, and as excavations at Lachish continue, it will be possible to obtain more data to determine broader trends in subsistence strategies and animal husbandry at Lachish.
References
Aharoni Y (1975) Investigations at Lachish: the sanctuary and the residency (Lachish V). Gateway Publishers, Tel Aviv
Baden-Powell D (1958) Shells. In: Tuffnel O (ed) Lachish IV: the Bronze Age. Oxford University Press, London, pp 323–324
Bar-Matthews M, Kaufman A (1988) Middle to late Holocene (6500 yr. period) paleoclimate in the Eastern Mediterranean region from stable isotopic composition of speleothems from Soreq Cave, Israel. In: Issar A, Brown N (eds) Water, Environment and Society in Times of Climatic Change. Kluwer Academic Press, Dordrecht, Boston, London, pp 203–214
Bartosiewicz, L., Van Neer, W., Lentacker, A., 1997. Draught cattle: their osteological identification and history.
Bar-Yosef Mayer D (2004) The mollusc shells. In: Ussishkin D (ed) The renewed archaeological excavations at Lachish (1973-1994)-V. Institute of Archaeology. Tel Aviv University, Tel Aviv, pp 2490–2503
Bate D (1953) The animal bones. In: Tuffnel O (ed) Lachish III: the Iron Age. Oxford University Press, London, pp 410–411
Bate D (1958) Animal bones. In: Tuffnel O (ed) Lachish IV: the Bronze Age. Oxford University Press, London, pp 322–323
Behrensmeyer AK (1978) Taphonomic and ecologic information from bone weathering. Paleobiology 4:150–162. https://doi.org/10.1017/S0094837300005820
Bietak M (2002) The function and some architectural roots of the Fosse Temple at Lachish. In: Ahituv S, Oren ED (eds) Aharon Kempinski Memorial Volume. Studies in Archaeology and Related Disciplines, Beer-Sheva Studies by the Department of Bible and Ancient Near East. Ben-Gurion University of the Negev Press, Beer-Sheva, pp 56–85
Boessneck J (1969) Osteological differences between sheep (Ovis aries Linné) and goat (Capra hircus Linné). In: Brothwell DR, Higgs ES (eds) Science in archaeology-a survey in progress and research. Thames and Hudson, London, London, pp 331–358
Burke A (2008) “Walled up to Heaven” the evolution of Middle Bronze Age fortification strategies in the Levant, studies in the Archaeology and History of the Levant. Eisenbrauns, Winona Lake, Indiana
Cochavi-Rainey, Z., Rainey, A.F., 2015. The El-Amarna correspondence: a new edition of the cuneiform letters from the site of El-Amamrna Based on Collations of All Extant Tablets. Brill.
Croft P (2004a) The osteological remains (Mammalian and Avian). In: Ussishkin D (ed) The renewed archaeological excavations at Lachish (1973-1994)-V. Institute of Archaeology. Tel Aviv University, Tel Aviv, pp 2254–2348
Croft P (2004b) Appendices I-VI. In: Ussishkin D (ed) The renewed archaeological excavations at Lachish (1973-1994)-V. Institute of Archaeology. Tel Aviv University, Tel Aviv, pp 2349–2455
Davis SJM (2000) The effect of castration and age on the development of the Shetland sheep skeleton and a metric comparison between bones of males, females and castrates. J Archaeol Sci 27:373–390. https://doi.org/10.1006/jasc.1999.0452
Davis SJ (2017) Towards a metrical distinction between sheep and goat astragali. In: Rowley-Conwy P, Serjeantson D, Halstead P (eds) Economic zooarchaeology. Studies in Hunting, Herding, and Early Agriculture. Oxbow Books, Oxford & Philadelphia, pp 50–82
Drori I, Horowitz A (1989) Tel Lachish: environment and subsistence during the Middle Bronze, Late Bronze and Iron Ages. Tel Aviv 16:206–211
Ducos P (1968) L’Origine des Animaux Domestiques en Palestine. Imprimeries Delmas, Bordeaux
Fisher JW (1995) Bone surface modifications in zooarchaeology. J Archaeol Method Theory 2:7–68
Garfinkel Y, Hasel MG, Klingbeil MG, Kang H-G, Choi G, Chang S-Y, Hong S, Ganor S, Kreimerman I, Ramsey CB (2019a) Lachish fortifications and state formation in the biblical kingdom of Judah in light of radiometric datings. Radiocarbon 61:695–712. https://doi.org/10.1017/RDC.2019.5
Garfinkel Y, Kreimerman I, Hasel MG, Klingbeil MG (2019b) First impression on the urban layout of the last Canaanite City of Lachish: a view from the Northeast Corner of the site. In: Maeir AM, Shai I, McKinny C (eds) The Late Bronze and Early Iron Ages of Southern Canaan. De Gruyter, Berlin, New York, pp 122–135
Golénischeff, W.S., 1913. Les papyrus hiératiques: no. 1115, 1116A et 1116B de l’Ermitage Impérial à St.-Pétersbourg. Manufacture des papiers de l’état.
Greenfield HJ, Arnold ER (2008) Absolute age and tooth eruption and wear sequences in sheep and goat: determining age-at-death in zooarchaeology using a modern control sample. J Archaeol Sci 35:836–849. https://doi.org/10.1016/j.jas.2007.06.003
Greenfield T, McKinny C, Shai I (2017) 18. I Can Count All My Bones: a preliminary report of the Late Bronze faunal remains from area B1 at Tel Burna, Israel. In: Lev-Tov J, Hesse P, Gilbert A (eds) The wide lens in archaeology: honoring Brian Hesse’s contributions to anthropological archaeology. Lockwood Press, Atlanta, pp 419–441
Habermehl K-H (1975) Altersbestimmung bei Haus-und Labortieren. P. Parey, Berlin, Hamburg
Kamlah J, Riehl S (in press) Atlas des Ackerbaus im antiken Palästina, RessourcenKulturen. Tübingen University Press, Tübingen
Klenck JD (2002) The Canaanite Cultic Milieu. In: The zooarchaeological evidence from Tel Haror, Israel, BAR international series. Archaeopress, Oxford
Koch I (2014) Goose keeping, elite emulation and Egyptianized feasting at Late Bronze Lachish. Tel Aviv 41:161–179. https://doi.org/10.1179/0334435514Z.00000000038
Langgut D, Finkelstein I, Litt T, Harald Neumann F, Stein M (2015) Vegetation and climate changes during the Bronze and Iron Ages (∼3600–600 BCE) in the Southern Levant Based on Palynological Records. Radiocarbon 57:217–235. https://doi.org/10.2458/azu_rc.57.18555
Lernau H (1975) Animal remains. In: Aharoni Y (ed) Investigations at Lachish: the sanctuary and the residency. Gateway, Tel Aviv, pp 86–103
Lernau H (1988) Mammalian Remains. In: ROthenberg B (ed) The Egyptian mining temple at Timna. Institute for Archaeo-Metallurgical Studies, London, pp 246–252
Lernau O, Golani D (2004) The osteological remains (Aquatic). In: Ussishkin D (ed) The renewed archaeological excavations at Lachish (1973-1994)-V. Institute of Archaeology. Tel Aviv University, Tel Aviv, pp 2456–2489
Lev-Tov, J.S.E., 2000. Pigs, philistines, and the ancient animal economy of Ekron from the Late Bronze Age to the Iron Age II.
Lev-Tov JSE (2010) A plebeian perspective on empire economies. Faunal remains from Tel Miqne-Ekron, Israel. In: Campana D, Crabtree P, DeFrance SD, Lev-Tov JSE, Choyke AM (eds) Anthropological approaches to zooarchaeology: colonialism, complexity and animal transformations. Oxbow, Oxford, pp 90–104
Lyman RL (1994) Vertebrate taphonomy. Cambridge University Press, Cambridge
Lyman RL (2008) Quantitative paleozoology, Cambridge Manuals in Archaeology. Cambridge University Press, Cambridge
Lyman R (2018) Observations on the history of zooarchaeological quantitative units: why NISP, then MNI, then NISP again? J Archaeol Sci Rep 18:43–50. https://doi.org/10.1016/j.jasrep.2017.12.051
Maher EF, Hesse B (2016) The Iron Age II faunal remains. In: Gitin S, Dothan T, Garfinkel Y (eds) Tel Miqne-Ekron Excavations, 1985–1988, 1990, 1992–1995: Field IV Lower—The Elite Zone, Part 3B: The Iron Age I and IIC Early and Late Philistine cities plans and sections. Eisenbrauns, Winona Lake, Indiana, pp 357–363
Maher EF, Hesse B (2016b) The Middle Bronze Age II and Iron Age I faunal remains. In: Gitin S, Dothan T, Garfinkel Y (eds) Tel Miqne-Ekron excavations. 1985–1988, 1990, 1992–1995: field IV Lower—The Elite Zone, Part 3B: The Iron Age I and IIC Early and Late Philistine Cities. Eisenbrauns, Indiana, pp 515–570
Mahler-Slasky Y, Kislev ME (2012) Preliminary archaeobotanical research at Tell es-Safi/Gath–The 1997-2002 Seasons. In: Maeir AM (ed) Tell Es-Safi/Gath I: The 1996–2005 Seasons, vol 1: Text. Harrassowitz, Wiesbaden, pp 579–587
Monchot H, Mashkour M, Vigne J-D (2005) Kernel smoothing and mixture analyses for the determination of the sex ratios at death, at the beginning of domestication of ungulates. In: Helmer D, Peters J, Vigne J-D (eds) The first steps of animal domestication: new archaeozoological approaches. Oxbow, Oxford, pp 55–60
Moran NC, O’Connor TP (1994) Age attribution in domestic sheep by skeletal and dental maturation: a pilot study of available sources. Int J Osteoarchaeol 4:267–285. https://doi.org/10.1002/oa.1390040402
Orendi A (2020) Flax cultivation in the southern Levant and its development during the Bronze and Iron Age. Quat Int 545:63–72
Payne S (1973) Kill-off patterns in sheep and goats: the mandibles from Aşvan Kale. Anatol Stud 23:281–303. https://doi.org/10.2307/3642547
Peck-Janssen SM (2006) Animal husbandry at Tell el Hesi (Israel): results from zooarchaeological and isotopic analysis. University of South Florida, Florida
Perevolotsky A, Perevolotsky A, Noy-Meir I (1989) Environmental adaptation and economic change in a pastoral mountain society: the case of the Jabaliyah Bedouin of the Mt. Sinai region Mt Res Dev 9:153–164
Redding R (1981) Decision making in subsistence herding of sheep and goats in the Middle East. University of Michigan, Michigan
Redding RW (1984) Theoretical determinants of a herder’s decisions: modeling variation in the sheep/goat ratio. Anim Archaeol 3:223–241
Riehl S, Shai I (2015) Supra-regional trade networks and the economic potential of Iron Age II sites in the southern Levant. J Archaeol Sci Rep 3:525–533. https://doi.org/10.1016/j.jasrep.2015.08.004
Rosen AM (1986) Environmental change and settlement at Tel Lachish. Israel Bull Am Sch Orient Res 263:55–60. https://doi.org/10.2307/1356910
Sapir-Hen L (2015) Faunal Remains from Tel Halif. In: Borowski O (ed) Faunal remains from Tel Halif. Emory University, Atlanta, Georgia, pp 167–181
Sass B, Garfinkel Y, Hasel MG, Klingbeil MG (2015) The Lachish jar sherd: an early alphabetic inscription discovered in 2014. Bull Am Sch Orient Res 374:233–245
Sasson A (2016) Animal husbandry in ancient Israel: a zooarchaeological perspective on livestock exploitation, herd management and economic strategies. Routledge, Oxon, New York
Simpson, E.H., 1949. Measurement of diversity. nature 163, 688–688.
Steele TE, Weaver TD (2002) The modified triangular graph: a refined method for comparing mortality profiles in archaeological samples. J Archaeol Sci 29:317–322. https://doi.org/10.1006/jasc.2001.0733
Stiner MC (1990) The use of mortality patterns in archaeological studies of hominid predatory adaptations. J Anthropol Archaeol 9:305–351
Stiner MC (1991) Food procurement and transport by human and non-human predators. J Archaeol Sci 18:455–482. https://doi.org/10.1016/0305-4403(91)90038-Q
Stiner MC (2005) The faunas of Hayonim Cave, Israel: a 200,000-year record of Paleolithic diet, demography, and society. Harvard University Press
Stiner MC, Kuhn SL, Weiner S, Bar-Yosef O (1995) Differential burning, recrystallization, and fragmentation of archaeological bone. J Archaeol Sci 22:223–237. https://doi.org/10.1006/jasc.1995.0024
Streit K, Webster L, Becker V, Jeske A-K, Misgav H, Höflmayer F (2018) Between destruction and diplomacy in Canaan: the Austrian-Israeli Expedition to Tel Lachish. East Archaeol 81:259–268
Tamar K, Bar-Oz G, Bunimovitz S, Lederman Z, Dayan T (2013) Geography and economic preferences as cultural markers in a border town: the faunal remains from Tel Beth-Shemesh, Israel: cultural identity: the faunal remains from Tel Beth-Shemesh. Israel Int J Osteoarchaeol 25:414–425. https://doi.org/10.1002/oa.2309
Tufnell O (1953) Lachish III: The Iron Age. Oxford University Press, London
Tufnell O (1958) Lachish IV (Tell Ed Duweir): The Wellcome-Marston Archaeological Expedition to the Near East. Oxford University Press, London, The Bronze Age
Tufnell O, Inge CH, Harding GL (1940) Lachish II-Tell Ed Duweir. Oxford University Press, London, The Fosse Temple
Ussishkin D (1977) The destruction of Lachish by Sennacherib and the Dating of the Royal Judean Storage Jars. Tel Aviv 4:28–60
Ussishkin, D., 1982. The conquest of Lachish by Sennacherib: an archaeological study of ancient Syro-Palestinian figurines. Tel Aviv Univ., Inst. of Archaeology.
Ussishkin, D., 2004. The renewed archaeological excavations at Lachish (1973-1994). Emery and Claire Yass Publications in Archaeology, Tel Aviv.
Ussishkin D (2014) Biblical Lachish: a tale of construction, destruction, excavation and restoration. Israel Exploration Society, Jerusalem
Von den Driesch A (1976) A guide to the measurement of animal bones from archaeological sites. Peabody Museum Press, Cambridge
Weaver TD, Boyko RH, Steele TE (2011) Cross-platform program for likelihood-based statistical comparisons of mortality profiles on a triangular graph. J Archaeol Sci 38:2420–2423. https://doi.org/10.1016/j.jas.2011.05.009
Webster L, Streit K, Dee MW, Hajdas I, Höflmayer F (2019) Identifying the Lachish of Papyrus Hermitage 1116A Verso and the Amarna Letters: implications of new radiocarbon dating. J Anc Egypt Interconnect 21:88–99
Weissbein I, Garfinkel Y, Hasel MG, Klingbeil MG (2016) Goddesses from Canaanite Lachish. Strata Bull Anglo-Isr Archaeol Soc 34
Weissbein I, Garfinkel Y, Hasel MG, Klingbeil MG, Brandl B, Misgav H (2019) The Level VI north-east temple at Tel Lachish. Levant 51:76–104
Zeder MA, Lapham HA (2010) Assessing the reliability of criteria used to identify postcranial bones in sheep, Ovis, and goats. Capra. J. Archaeol. Sci. 37:2887–2905. https://doi.org/10.1016/j.jas.2010.06.032
Zeder MA, Pilaar SE (2010) Assessing the reliability of criteria used to identify mandibles and mandibular teeth in sheep, Ovis, and goats. Capra J Archaeol Sci 37:225–242. https://doi.org/10.1016/j.jas.2009.10.002
Acknowledgements
We would like to thank Adriano Orsingher and Effrosyni Roditi for helping us enter data in our faunal database and check the chronology of the previous faunal reports. Thanks, too, to Marco Nicolì for his insights on early drafts of this paper. Thanks to the Lachish excavation team, without whom this study would not have been possible. The excavations at Lachish are part of the project ‘Tracing Transformations’ directed by Felix Höflmayer and funded by the Austrian Science Fund (FWF START Y 932-G25). We also thank Christoph Forster (Datalino, Berlin) for making the map featuring Lachish and the comparative sites. Finally, we thank the anonymous reviewers for their comments which helped improve this article.
Code availability
Not applicable
Funding
Open Access funding enabled and organized by Projekt DEAL. The research conducted here was made possible by funding from the CRC 1070 ResourceCultures, project A05 at the University of Tübingen.
Author information
Authors and Affiliations
Contributions
Shyama Vermeersch: conceptualization, formal analysis, writing—original draft, visualization. Simone Riehl: writing—review and editing. Britt M. Starkovich: writing–review and editing. Katharina Streit: resources, writing—review and editing. Felix Höflmayer: resources, writing—review and editing
Corresponding author
Ethics declarations
Ethics approval
Not applicable
Consent to participate
Not applicable
Consent for publication (include appropriate statements)
All authors agree with the content of this manuscript and approve of it being published.
Conflict of interest
The authors declare no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Vermeersch, S., Riehl, S., Starkovich, B.M. et al. Animal husbandry from the Middle Bronze Age through the Iron Age in the Shephelah—faunal remains from the new excavations at Lachish. Archaeol Anthropol Sci 13, 38 (2021). https://doi.org/10.1007/s12520-021-01289-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-021-01289-1