
Abstract
Salt stress affects the growth and development of plants, which results in a decrease in crop quality and yield. In this study, we used tomato seedlings treated with salt and trehalose as experimental materials and analyzed them using the technique for order preference by similarity to ideal solution analysis to select the optimal trehalose concentration for treatment. We also determined the contents of sugar and abscisic acid (ABA) and detected the expression of genes involved in the metabolism of sugar and ABA by quantitative real-time PCR. Results showed that the optimal trehalose concentration was 2 mmol/L for tomato seedlings under salt stress. Exogenous trehalose decreased the starch content and increased the soluble sugar content by affecting the expression of genes related to the metabolism of starch and soluble sugar. Exogenous trehalose altered the accumulation and distribution of sugar by inducing the upregulation of sugar transporter genes. Furthermore, trehalose increased the ABA content to induce salt stress response by regulating the expression of genes related to the synthesis and metabolism of ABA. In conclusion, trehalose can effectively alleviate salt stress and enhance salt tolerance of tomato. These findings provide a novel perspective and a better resource to investigate the salt tolerance mechanism and a new method for alleviating salt stress in tomato.
Similar content being viewed by others


Introduction
In recent years, increasing research attention has been paid to salt stress that can affect the growth and development of plants, especially in the stage of seed germination [1, 2]. The response of plants to salt stress is complex, which is achieved through a variety of physiological and biochemical reactions and molecular mechanisms, and also involves the differential expression of stress-related genes in the corresponding pathways [3,4,5].
Soluble sugars as osmoprotectants help in the alleviation of the negative effects of salt on plants. Rosa et al. [6] suggested that there was a significant increase in the levels of glucose, sucrose, and fructose in plants under salt stress, which played a vital role in carbon storage, osmotic protection, and free radical scavenging. Nemati et al. [7] demonstrated that exogenous glucose can limit the accumulation of Na+ and stimulate the uptake of K+, thereby maintaining ion homeostasis under salt stress.
Trehalose was first isolated from ryegrass by Wiggers in the late nineteenth century and was later found in bacteria, algae, yeast, lower plants, insects, and other invertebrates [8]. The amount of trehalose present in plants is extremely low, and excessive accumulation of trehalose can affect the metabolism of normal carbohydrates in plants and even inhibit their growth [9,10,11]. Trehalose produces a marked effect primarily in the form of signaling molecules under stress conditions [12]. The metabolic pathway of trehalose in plants is recognized as the Ots A–Ots B pathway, in which under the action of trehalose phosphate synthase, uridine diphosphate glucose and glucose-6-phosphate form trehalose-6-phosphate and uridine diphosphate and then generate trehalose and inorganic phosphates under the catalysis of trehalose phosphatase, as shown in Fig. 1 [13, 14]. Garcia et al. [15] reported in 1997 that when rice was exposed to salt stress, the addition of low concentrations of trehalose (1–10 mmol/L) could protect its root integrity, protect root cells from abnormal cell division induced by severe salt stress, and excrete excessive Na+ from cells by protecting ion pumps. Yang et al. [16] demonstrated that trehalose can increase the salt tolerance of plants, while exogenous trehalose may act as an inducer of biological processes involved in the salt stress response of plant.

Metabolic pathway of trehalose in plants
Sugars not only participate in the metabolism of carbon and energy in cells but also play a key role in plant growth and stress resistance as a signal molecule [17]. The presence of an interaction has been suggested between sugars and plant hormones in metabolic pathways and stress response [18, 19]. Cheng et al. [20] found that exogenous glucose enhanced the expression of genes related to the biosynthesis of abscisic acid (ABA) and ultimately increased the ABA content in plants.
However, the effects of exogenous trehalose on sugar metabolism and the interaction between trehalose and ABA under salt stress are still unclear. In previous studies, we used transcriptome sequencing to screen the pathways and the related genes involved in protein metabolism and carbohydrate metabolism pathways in tomato [21]. In response to salt stress, the protein metabolism pathway was significantly enriched in seven pathways, with 17 key genes being differentially expressed, and the carbohydrate metabolism pathway was significantly enriched in six pathways, with 19 key genes being differentially expressed. Based on transcriptome sequencing results, this study analyzed the metabolic changes in sugar and ABA in tomato seedlings treated with trehalose and salt and determined the differential expression profiles of key genes involved in sugar metabolism. The purpose of this study was to investigate the effects of trehalose on salt stress and the interaction between sugar metabolism and trehalose on regulating the response to salt stress in tomato, which could provide a theoretical basis for improving crop yield, salt tolerance of plants, and the utilization rate of saline–alkali land in agricultural research.
Materials and Methods
Plant Material and Treatment
After soaking for 4–6 h at room temperature, tomato (cv. Ailsa Craig) seeds were sterilized with 75% ethanol and 10% NaClO, cultured at 25 °C for 8 d, and then cultured in Hoagland’s nutrient solution for 4 weeks with illumination for 16 h/d at 25 °C. Trehalose and salt were applied to tomato seedlings with consistent growth, and each treatment was performed for 7 d as follows: (T1) control: 0 mmol/L Tre + 0 mmol/L NaCl; (T2) 0 mmol/L Tre + 200 mmol/L NaCl; (T3) 1 mmol/L Tre + 200 mmol/L NaCl; (T4) 2 mmol/L Tre + 200 mmol/L NaCl; (T5) 3 mmol/L Tre + 200 mmol/L NaCl; (T6) 5 mmol/L Tre + 200 mmol/L NaCl; and (T7) 10 mmol/L Tre + 200 mmol/L NaCl. Salt concentration was selected based on our previous research [21]. The formulation of Hoagland’s nutrient solution was as follows: pH 6.0, 945 mg/L Ca(NO3)2, 607 mg/L KNO3, 115 mg/L (NH4)3PO4, 493 mg/L MgSO4, 2.5 mL/L ferric salt solution, and 5 mL/L trace element.
Determination of Growth Indicators and Related Physiological Indicators
Plant height, fresh mass, and dry mass of tomato seedlings were measured, and the relative water content was calculated as follows:
The chlorophyll content of tomato leaves was measured by the acetone method as previously described in Ref. [22] with slight modification. The activity of peroxidase (POD) was determined by the guaiacol method, and the activity of superoxide dismutase (SOD) was determined by the photochemical reaction method of nitroblue tetrazolium according to previously described methods, with slight modification [23, 24]. Catalase kit was used to determine the activity of CAT enzyme according to the instructions (Solarbio, Beijing, China). The level of proline was determined as described in Ref. [25].
For the determination of malondialdehyde (MDA) levels, 1.0 g seedlings was added to 10 mL of 10% trichloroacetic acid solution, ground on ice, and then centrifuged at 5000 g. Then, 1 mL supernatant was aspirated and 3 mL of 0.6% thiobarbituric acid solution was added and heated in boiling water for 15 min. After cooling the liquid rapidly, absorbance values at 450, 532, and 600 nm were measured. The MDA level was determined according to the following formula:
The optimum trehalose treatment concentration was selected using the technique for order preference by similarity to ideal solution (TOPSIS) analysis.
Determination of Starch and Soluble Sugar Contents Under Optimal Trehalose Concentration
Tomato seedlings (cultivated as mentioned earlier) with consistent growth were selected for four treatments (0 mmol/L Tre + 0 mmol/L NaCl; 0 mmol/L Tre + 200 mmol/L NaCl; 2 mmol/L Tre + 0 mmol/L NaCl; and 2 mmol/L Tre + 200 mmol/L NaCl), and each treatment was processed separately for 0, 1, 3, 5, and 7 d. Both roots and leaves of tomato were selected for determination.
Starch and soluble sugar contents were evaluated based on a previously reported protocol, with some modifications [25].
For the determination of trehalose content, 2 g tomato seedlings were ground into powder in a mortar with liquid nitrogen and homogenized by adding 2 mL of 80% ethanol and 1% polyvinylpolypyrrolidone (PVPP). The supernatant was concentrated in 200 mL distilled water and filtered by a microporous membrane (0.45 μm). Trehalose content was determined using the trehalose assay kit (Seebio, China).
Determination of the Genes Expression Related to Sugar Metabolism Under Optimal Trehalose Concentration
Total RNA in tomato leaves and roots was extracted using the Column Plant RNAOUT kit (Tiandz, Beijing, China), and cDNA was synthesized following the instruction of the FastQuant RT kit (Tiangen Biotech, Beijing, China). CDS sequences of internal reference gene and target genes were searched in the NCBI database. The Primer 5.0 software was used to design specific primer sequences given in Table 1. PCR amplification and quantitative real-time PCR (qRT-PCR) were performed using S1000 thermal cycler (BIO-RAD, USA) and LightCycler96 (Roche, Basel, Switzerland), respectively. β-Tublin was selected as the internal reference gene [26].
Determination of the Genes Expression Related to Sugar Transporter Under Optimal Trehalose Concentration
The expression of genes related to sugar transporter was measured as mentioned earlier. The primer sequences are given in Table 2.
Determination of ABA Content and the Expression of Metabolism-Related Genes Under Optimal Trehalose Concentration
After being ground with liquid nitrogen, 0.1 g of tomato seedlings was added with 0.8 mL of 10% methanol and 1% acetic acid, and then extracted at 4 °C for 16 h. After centrifuged at 10000g for 15 min, the supernatant was aspirated and filtered through a 0.45-μm microporous membrane. The ABA content was determined using a Plant ABA ELISA test kit (Jianglai Biotechnology, Shanghai, China).
The expression of metabolism-related genes was determined as mentioned earlier. The primer sequences are given in Table 3.
Statistical Analysis
All experiments were repeated three times. Significance of differences (p < 0.05) between the mean values was determined by analysis of variance using the IBM SPSS Statistics 20 software (SPSS commercial software, USA).
Results
Screening for Optimal Trehalose Concentration
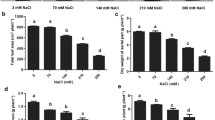
Under salt stress, the tomato leaves exhibited a withering state, and the plant height, fresh mass, and the relative water content (RWC) of the leaves were significantly reduced as shown in Figs. 2 and 3, respectively. However, exogenous trehalose improved the growth of tomato and increased the plant height, fresh mass, and the relative water content of tomato leaves, especially under treatment with 2 mmol/L trehalose, which was consistent with the results in Ref. [27].

Tomato leaves grown under normal and salt-treated conditions

Effect of trehalose concentration on tomato growth under salt stress. A Plant height, B fresh mass, and C relative water content
Salt stress affects photosynthesis in plants. Salt stress can increase the number of free radicals in chloroplasts, thereby destroy the chlorophyll, and affect the photosynthesis of plants. After exogenous trehalose treatment, the reduction in chlorophyll content was improved and plant salt stress was alleviated [28]. The chlorophyll content in leaves decreased significantly after salt stress treatment (Fig. 4). However, after treatment with different concentrations of exogenous trehalose, the chlorophyll content increased initially and then decreased. The highest chlorophyll content was found to be with 2 mmol/L trehalose treatment.

Effect of trehalose concentration on the chlorophyll content of tomato leaves under salt stress. A Chlorophyll a, B chlorophyll b, and C total chlorophyll
The MDA content increased significantly after salt treatment as shown in Fig. 5A, indicating that salt stress destroyed the cell membrane of plants severely, and a large number of free radicals and reactive oxygen species were produced that resulted in membrane damage. After treatment with different concentrations of trehalose, the MDA content decreased initially and then increased and was significantly reduced with 2 mmol/L trehalose treatment.

Effects of different concentrations of trehalose on physiological indexes of tomato leaves under salt stress. A MDA, B proline, C CAT, D SOD, and E POD
The accumulation of proline increases the osmotic potential of cells, regulates osmosis, and alleviates salt stress. Under salt treatment, proline showed a certain accumulation; after the application of trehalose, the proline content increased significantly, with the highest content being observed with 2 mmol/L trehalose treatment (Fig. 5B).
High salt content causes an imbalance of reactive oxygen species in plants, and plants remove the excess reactive oxygen species primarily through antioxidant enzymes such as SOD, POD, CAT, and other antioxidant enzymes and non-antioxidant enzymes such as ascorbic acid, glutathione (GSH), β-carotene, and mannitol [21]. It has been reported that the activities of SOD, CAT, and POD show a positive correlation with plant salt resistance [29]. In our study, the activities of these three antioxidant enzymes (POD, SOD, and CAT) increased after salt treatment, which indicated that salt stress could enhance the activities of these enzymes. After treatment with different concentrations of trehalose, the activities of the three antioxidant enzymes increased initially and then decreased, with the highest activity of CAT and SOD being detected with 2 mmol/L trehalose treatment (Fig. 5C, D). And the activity of POD reached the second highest, close to the highest, with 2 mmol/L trehalose treatment (Fig. 5E).
The TOPSIS mathematical model was used to analyze and screen the optimum exogenous trehalose concentration for alleviating salt stress in tomato seedlings. Results showed that the optimum concentration of trehalose was 2 mmol/L (Table 4). Therefore, this concentration was selected for subsequent experiments.
Effect of Exogenous Trehalose on Sugar Content of Tomato Seedlings Under Salt Stress
The accumulation of starch can lead to a negative feedback mechanism of photosynthesis, reduce the intensity of photosynthesis, and slow down plant growth. Under salt stress, starch accumulated in tomato leaves and reached the maximum content on the 7th day, whereas the addition of trehalose reduced starch accumulation, as shown in Fig. 6. The changes in the starch content of roots were consistent with those of leaves, but the starch content in roots was lower than that in leaves, which may be due to the fact that salt stress destroyed the transport mechanism of starch from leaves to roots and affected starch transport, thus primarily accumulating in leaves and causing a negative feedback mechanism of photosynthesis.

Effect of exogenous trehalose on the starch content of tomato seedlings under salt stress. A Roots and B leaves
Plants regulate the cell osmotic potential by absorbing external inorganic salt ions and synthesizing small organic molecules such as proline and soluble sugar (sucrose, trehalose, etc.) to resist salt stress [30, 31]. Soluble sugar can be used as an osmotic substance to alleviate the ionic stress of plants and as a signal molecule to stimulate the response to salt stress. Our study showed that under salt stress, the soluble sugar contents increased in tomato roots and leaves, and exogenous trehalose significantly increased the soluble sugar contents, especially in the later stage (Fig. 7). It may be due to the fact that trehalose reduced the accumulation of starch and promoted the conversion of starch into soluble sugar.

Effect of exogenous trehalose on the soluble sugar content of tomato seedlings under salt stress. A Roots and B leaves
As shown in Fig. 8, exogenous trehalose treatment under salt stress increased sucrose content, which is the primary product of photosynthesis. Exogenous trehalose contributed to the accumulation of sucrose, indicating that it may induce an increase in photosynthesis rate, which was consistent with the abovementioned results showing that exogenous trehalose increased the content of photosynthetic pigments. The change in fructose levels was inconsistent compared with sucrose levels. Under salt stress, the fructose content increased, whereas exogenous trehalose treatment significantly decreased the fructose content. It was probably because trehalose reduced the hydrolysis rate of sucrose, and the rate of sucrose synthesis was greater than the rate of degradation. With the progression of salt treatment, the sorbitol content increased gradually. Exogenous trehalose further induced an increase in sorbitol content in the later stage.

Effects of exogenous trehalose on the contents of sucrose, fructose, and sorbitol in tomato seedlings under salt stress. A Sucrose in roots, B sucrose in leaves, C fructose in roots, D fructose in leaves, E sorbitol in roots, and F sorbitol in leaves
In general, the content of trehalose is extremely low in plants, and it is difficult to detect its presence directly. As shown in Fig. 9, the trehalose content under normal conditions was very low, and salt stress stimulated trehalose production, especially in leaves. The increase in trehalose content confirmed that it enhanced the salt tolerance of tomato to some extent.

Effect of exogenous trehalose on the trehalose content of tomato seedlings under salt stress. A Roots and B leaves
Effect of Exogenous Trehalose on Key Genes Involved in Sugar Metabolism in Tomato Seedlings Under Salt Stress
Starch is the primary product of plant photosynthesis and is generally degraded to resist stress. The starch synthase (SS) gene and the adenosine diphosphate glucose pyrophosphate (AGPase) gene play an important role in the starch synthesis pathway, while alpha amylase (AMY) and beta amylase (BAM) are the two key enzymes involved in plant starch degradation.
In our study, SS expression in tomato roots decreased initially and then increased under salt stress (Fig. 10). On the 1st day, exogenous trehalose significantly inhibited the expression of SS, which was consistent with the decrease in starch content. However, with the progression of salt treatment, SS expression gradually became stable. The change in the expression of SS in tomato leaves was approximately similar to that in roots. Under salt stress, AGPase expression in tomato roots was inhibited, especially in the case of exogenous trehalose treatment, which decreased its expression to 22.3% on the 5th day. However, in leaves, AGPase expression was increased significantly on the 1st day and then decreased with the progression of salt treatment.

Effect of exogenous trehalose on the expression of SS and AGPase in tomato seedlings under salt stress. ASS in roots, BSS in leaves, CAGPase in roots, and DAGPase in leaves
As shown in Fig. 11, beta amylase played an important role in response to salt stress. The expression of the gene BAM in tomato roots was significantly upregulated compared with the control group, especially under treatment with exogenous trehalose (except for the 5th day), which promoted the degradation of starch and reduced the feedback inhibition of photosynthesis. In leaves, BAM expression increased initially and then decreased, which was basically consistent with the previous changes in starch content. Salt + trehalose treatment significantly upregulated the expression of the α-amylase gene AMY, with the highest expression being observed on the 3rd day in roots, which was approximately eight times of that in the control group and two times of that in the group of salt treatment alone, indicating that AMY was sensitive to trehalose. In leaves, the expression of AMY reached the peak on the 1st day and then decreased. The differential expression of AMY in different tissues indicated that its contribution to starch degradation was inconsistent and tissue specific.

Effect of exogenous trehalose on the expression of BAM and AMY in tomato seedlings under salt stress. ABAM in roots, BBAM in leaves, CAMY in roots, and DAMY in leaves
Sucrose constitutes the highest proportion of sugars in plants and is directly related to plant stress resistance. The sucrose phosphate synthase (SPS) gene, the sucrose synthase (SUS3) gene, and the acid invertase (Wiv-1) gene are important in the sucrose metabolism pathway.
Figure 12 shows that SPS expression increased with the addition of exogenous trehalose under salt stress. Especially in roots, SPS expression was 14 times higher than that in the control group after 1 day of treatment, indicating that the effect of exogenous trehalose on roots was more obvious, which was consistent with the theory that roots are the most sensitive organs under salt stress. Sucrose synthase not only hydrolyzes sucrose but also synthesizes sucrose, regulating a reversible reaction. In roots, the expression of SUS3 was not significantly different from that in the control group and tended to be stable. In leaves, SUS3 expression decreased initially and then increased compared with that in the control group.

Effect of exogenous trehalose on the expressions of SPS, SUS3, and Wiv-1 in tomato seedlings under salt stress. ASPS in roots, BSPS in leaves, CSUS3 in roots, DSUS3 in leaves, EWiv-1 in roots, and FWiv-1 in leaves
The enzyme acid invertase facilitates the hydrolysis of sucrose into glucose and fructose, which can significantly increase the cell osmotic potential. Salt treatment induced the expression of Wiv-1 in roots and leaves. Especially on the 3rd day, Wiv-1 expression reached the peak with salt + trehalose treatment, which was upregulated by 7.26 times and 10.68 times, respectively, indicating that Wiv-1 may play an essential role in salt stress, while exogenous trehalose treatment significantly stimulated its expression.
The trehalose phosphate synthase (TPS) gene, the trehalose phosphatase (TPP) gene, and the trehalose (Tre) gene play an important role in the synthesis and metabolism of trehalose. As shown in Fig. 13, salt stress induced the expression of the TPS gene in roots and leaves, and the upregulation was higher with salt + trehalose treatment, which was consistent with the trend of trehalose content in Fig. 9. The expression of TPP was significantly upregulated under salt stress, reaching peaks on the 3rd and 1st day in roots and leaves, respectively. Exogenous trehalose treatment inhibited the expression of this gene, which may be caused due to the negative feedback regulation. The expression of Tre in roots and leaves increased initially and then decreased and was enhanced by the addition of trehalose and yielded a peak after 1 day.

Effect of exogenous trehalose on the expression of TPS, TPP, and Tre in tomato seedlings under salt stress. ATPS in roots, BTPS in leaves, CTPP in roots, DTPP in leaves, ETre in roots, and FTre in leaves
Effects of Exogenous Trehalose on Sugar Signal Transduction Genes and Sugar Transporter Genes of Tomato Seedlings Under Salt Stress
Sucrose is converted into fructose under the action of sucrose synthase and hydrolase, and fructose enters the glycolysis cycle under the action of fructose phosphokinase and phosphokinase, participating in plant stress response. Under salt stress, the expression of fructokinase (FRK2) gene decreased initially and then increased in roots, whereas salt + trehalose treatment significantly inhibited its expression, which began to be upregulated on the 7th day (Fig. 14). In leaves, the expression of FRK2 increased gradually under salt + trehalose treatment, reaching its peak on the 5th day.

Effect of exogenous trehalose on the expression of FRK2 and PFK in tomato seedlings under salt stress. AFRK2 in roots, BFRK2 in leaves, CPFK in roots, and DPFK in leaves
As shown in Fig. 14C, D, salt stress significantly induced the expression of the phosphate fructokinase (PFK) gene, indicating that PFK may play an important role in response to salt stress. Moreover, salt + trehalose treatment further increased PFK expression in roots, which was upregulated by 9.3 times compared with that in the control group on the 1st day. PFK expression in tomato leaves was lower than that in roots but significantly upregulated.
Sucrose and hexose signal receptors are the major sugar signal receptors in plants. We selected two hexokinase genes, HXK2 and HXK3, for this study (Fig. 15). Both genes were upregulated to varying degrees under salt stress, and exogenous trehalose was more conducive to the expression of hexokinase gene. Under salt + trehalose treatment, the expression of HXK2 increased initially and then decreased, reaching its peak on the 3rd day. HXK3 expression was upregulated with both salt treatment and salt + trehalose treatment.

Effect of exogenous trehalose on the expression of HXK2 and HXK3 in tomato seedlings under salt stress. AHXK2 in roots, BHXK2 in leaves, CHXK3 in roots, and DHXK3 in leaves
The transport and distribution of sugar directly affect the normal growth and development of plants and also play an important role in plant stress. We analyzed the changes in the expression of the monosaccharide transporter (MST3) gene and the sucrose transporter genes (SUT1 and SUT4) under salt stress. As shown in Fig. 16, the expression of MST3 in roots was upregulated with salt treatment and significantly upregulated with salt + trehalose treatment. MST3 expression in leaves was also induced by salt stress and reached the peak on the 1st day after salt + trehalose treatment. These results showed that both SUT1 and SUT4 were upregulated with salt + trehalose treatment, suggesting that these genes may be involved in the response to salt stress.

Effect of exogenous trehalose on the expression of MST3, SUT1, and SUT4 in tomato seedlings under salt stress. AMST3 in roots, BMST3 in leaves, CSUT1 in roots, DSUT1 in leaves, ESUT4 in roots, and FSUT4 in leaves
Effects of Exogenous Trehalose on ABA Synthesis and Metabolism of Tomato Seedlings Under Salt Stress
Under stress conditions, there is an increase in the ABA content in plants to resist external stress. As shown in Fig. 17, the ABA content increased significantly under salt stress, indicating that ABA was a stress-related hormone. After salt treatment, exogenous trehalose significantly increased the ABA content in tomato roots and leaves, reaching their peaks on the 5th day.

Effect of exogenous trehalose on the ABA content in tomato seedlings under salt stress. A Roots and B leaves
NCED1 and NCED2 are key genes in the ABA synthesis pathway. As shown in Fig. 18, NCED1 and NCED2 in tomato roots and leaves were upregulated under salt treatment, especially when exogenous trehalose was added, indicating that exogenous trehalose had a significant positive effect on ABA synthesis.

Effect of exogenous trehalose on the expression of NCED1 and NCED2 in tomato seedlings under salt stress. ANCED1 in roots, BNCED1 in leaves, CNCED2 in roots, and DNCED2 in leaves
CYP707A1 and CYP707A2 are important genes in the ABA metabolic pathway. Figure 19 shows that the expression of CYP707A1 was upregulated under salt stress, especially in roots. When treated for 3 days, CYP707A1 expression was 1.62 times that of the control group. However, the addition of exogenous trehalose reduced the upregulated expression of CYP707A1 under salt stress. CYP707A2 expression was decreased, and exogenous trehalose further reduced its expression.

Effect of exogenous trehalose on the expression of CYP707A1 and CYP707A2 in tomato seedlings under salt stress. ACYP707A1 in roots, BCYP707A1 in leaves, CCYP707A2 in roots, and DCYP707A2 in leaves
Discussion
Salt treatment inhibited the growth of tomato seedlings. After the application of trehalose, the plant damage was alleviated, whereas high concentrations of trehalose inhibited the growth of tomato seedlings. Although trehalose can provide a source of carbon for tomato, due to the inability of tomato to utilize carbon source under salt stress, ionic stress is aggravated, which reduces photosynthesis and affects tomato growth.
ROS accumulation increases significantly under salt stress, which may lead to cell membrane damage, oxidative stress, or cell death [32, 33]. In this study, we demonstrated that trehalose exerted the effects of enhancing the activities of antioxidant enzymes, increasing the proline content, and decreasing the MDA content, therefore alleviating the damage caused due to salt stress.
Under salt stress, starch accumulated in tomato roots and leaves, leading to a negative feedback mechanism of photosynthesis, which reduced the photosynthetic rate, decreased the chlorophyll content, and hindered starch transport. Exogenous trehalose treatment inhibited the expression of the starch synthesis genes SS and AGPase and promoted the expression of the starch metabolism genes BAM and AMY, thereby reducing the starch content, alleviating the negative feedback mechanism, and facilitating the accumulation of photoassimilation products.
Soluble sugars are primarily used as osmotic agents to prevent plant cells from dehydration and death and to maintain the internal stability of the cells under salt stress [34, 35]. When treated with salt, the content of various soluble sugars in different tissues of tomato seedlings increased significantly, while exogenous trehalose further increased their contents, especially that of sucrose. As an important osmotic regulator, sucrose can significantly increase the osmotic potential of plant cells and alleviate ionic stress. In addition, sucrose can eliminate ROS in plants. Exogenous trehalose administration significantly induced the expression of the SPS under salt stress and increased the sucrose content in tomato. The expression of the sucrose synthase gene SUS3 increased initially and then decreased, whereas that of the acid invertase gene Wiv-1 increased, which maintained a relative stability of sucrose content under salt stress.
The expression of the TPS in tomato was upregulated under salt stress, which was consistent with the findings of Li et al. [36] who showed that overexpression of OsTPS1 enhanced the salt tolerance of rice and exogenous trehalose treatment further increased TPS expression. In our experiment, exogenous trehalose administration increased the content of endogenous trehalose in tomato, resulted in a negative feedback regulation, inhibited the expression of the TPP, enhanced the expression of trehalose gene, and promoted the conversion of excessive trehalose into glucose to maintain the stability of its content. TPS, TPP, and Tre exhibited differential expressions under salt stress, suggesting that the trehalose metabolic pathway could directly affect the salt tolerance of plants.
Fructose phosphokinase and fructokinase are two important enzymes in the plant glycolytic pathway, which can maintain the normal physiological function of plants and alleviate the physiological damage occurring in plants under stress [37]. The results of this study showed that the addition of exogenous trehalose could significantly upregulate the expressions of FRK and PFK under salt stress, suggesting that it could help plants accumulate sugars and maintain strong respiration. Sugar not only is the primary energy source for plant growth, but also can be used as a signal molecule to respond to plant stress. HXK is a sugar receptor that is affected by the external environment and forms a network signal system with plant hormones such as ethylene, ABA, and gibberellin. Exogenous trehalose treatment induced the upregulation of HXK2 and HXK3 to different degrees under salt stress. Sami et al. [38] reported that sugar and ABA interacted in the metabolic pathways of plants to influence drought and salt resistance.
Salt stress affects the activities of sugar transporters on plant membranes, which results in the abnormal transport of sugar and the inability of plants to obtain energy [39]. The sugar transporter genes exhibited different expression trends during osmotic stress, suggesting that the source–sink relationship of carbon may be related to the regulation of sugar transporter gene expression [40]. In addition, AtSUC4 could regulate sucrose distribution and metabolism in response to drought stress [41]. The results of this study demonstrated that the expressions of the monosaccharide transporter (MST3) gene and the sucrose transporter genes SUT1 and SUT4 were significantly upregulated with salt + trehalose treatment. Trehalose may change the source–sink transport pathway of sugars, thereby changing the accumulation and distribution of sugars to respond to salt stress.
The entire process of plant growth and development is affected by the interaction between sugar and ABA. In this experiment, the ABA content of tomato seedlings accumulated significantly under salt stress, which was consistent with the results of Kong et al. [42] and Shu et al. [43], indicating that ABA directly responded to plant salt stress. Salt + trehalose treatment resulted in the upregulation of the ABA synthesis-related genes and the downregulation of metabolic genes, which indicated that the tomato plant increased its ABA content by accelerating ABA synthesis and reducing ABA decomposition. Saeedipour [44] found that differences in salt stress tolerance of rice plant were related to its ABA synthesis ability under stress conditions. Tomatoes overexpressing NCED accumulated more ABA and showed improved drought resistance. ABA primarily improves plant stress resistance by closing the stomata of plant leaves, reducing transpiration, and maintaining water in plants [45]. Therefore, trehalose acts as a signaling molecule by affecting the ABA anabolic pathway, leading to ABA accumulation and further improving the salt tolerance of tomato.
In summary, trehalose, a soluble, non-reducing disaccharide, is present in a low amount in plants; however, it regulates sugar accumulation and distribution by affecting the activities of sugar transporters and simultaneously regulates sugar metabolism and ABA metabolism, thereby significantly improving plant stress resistance (Figs. 20, 21). This study provides a greater understanding of the salt-tolerant mechanism of plants and has important value for improving crop yield, salt tolerance of plants, and utilization of saline–alkali land in agricultural research.

Changes in glucose metabolism in tomato seedlings under salt stress by exogenous trehalose

Changes in ABA metabolism in tomato seedlings under salt stress by exogenous trehalose
Future research will continue to explore the effect of trehalose on tomato salt stress using transgenic technology to alter the expression of genes related to trehalose metabolism, which could help in further studying the salt tolerance mechanism of trehalose.
References
Cheng Y, Zhang XX, Sun TY et al (2018) Glutamate receptor homolog3.4 is involved in regulation of seed germination under salt stress in Arabidopsis. Plant Cell Physiol 59(5):978–988
Liu H, Chen XL, Song L et al (2019) Polysaccharides from Grateloupia filicina enhance tolerance of rice seeds (Oryza sativa L.) under salt stress. Int J Biol Macromol 124:1197–1204
Espasandin FD, Calzadilla PI, Maiale SJ et al (2018) Overexpression of the arginine decarboxylase gene improves tolerance to salt stress in Lotus tenuis plants. J Plant Growth Regul 37(1):156–165
Lu P, Magwanga R, Lu HJ et al (2018) A novel G-protein-coupled receptors gene from upland Cotton enhances salt stress tolerance in transgenic Arabidopsis. Genes 9(4):209
Denver JB, Ullah H (2019) MiR393 s regulate salt stress response pathway in Arabidopsis thaliana through scaffold protein RACK1A mediated ABA signaling pathways. Plant Signal Behav 14(6):1600394. https://doi.org/10.1080/15592324.2019.1600394
Rosa M, Prado C, Podazza G et al (2009) Soluble sugars: metabolism, sensing and abiotic stress: a complex network in the life of plants. Plant Signal Behav 4(5):388–393
Nemati I, Moradi F, Gholizadeh S et al (2011) The effect of salinity stress on ions and soluble sugars distribution in leaves, leaf sheaths and roots of rice (Oryza sativa L) seedlings. Plant Soil Environ 57(1):26–33
Richards AB, Krakowka S, Dexter LB et al (2002) Trehalose: a review of properties, history of use and human tolerance, and results of multiple safety studies. Food Chem Toxicol 40(7):871–898
Paul MJ, Gonzalez-Uriarte A, Griffiths CA et al (2018) The role of trehalose 6-phosphate in crop yield and resilience. Plant Physiol 177(1):12–23
Luo Y, Wang W, Fan YZ et al (2018) Exogenously-supplied trehalose provides better protection for D1 protein in winter wheat under heat stress. Russ J Plant Physiol 65(1):115–122. https://doi.org/10.1134/s1021443718010168
Vogel G, Fiehn O, Jean-Richard-dit-Bressel L et al (2001) Trehalose metabolism in Arabidopsis: occurrence of trehalose and molecular cloning and characterization of trehalose-6-phosphate synthase homologues. J Exp Bot 52(362):1817–1826. https://doi.org/10.1093/jexbot/52.362.1817
Paul MJ (2008) Trehalose 6-phosphate: a signal of sucrose status. Biochem J 412(1):e1–e2
Voit EO (2003) Biochemical and genomic regulation of the trehalose cycle in yeast: review of observations and canonical model analysis. J Theor Biol 223(1):55–78
Penna S (2003) Building stress tolerance through over-producing trehalose in transgenic plants. Trends Plant Sci 8(8):355–357
Garcia AB, Engler J, Iyer S et al (1997) Effects of osmoprotectants upon NaCl stress in rice. Plant Physiol 115(1):159–169
Yang L, Zhao XJ, Zhu H et al (2014) Exogenous trehalose largely alleviates ionic unbalance, ROS burst, and PCD occurrence induced by high salinity in Arabidopsis seedlings. Front Plant Sci 5:570. https://doi.org/10.3389/fpls.2014.00570
Gazzarrini S, McCourt P (2001) Genetic interactions between ABA, ethylene and sugar signaling pathways. Curr Opin Plant Biol 4(5):387–391. https://doi.org/10.1016/s1369-5266(00)00190-4
Finkelstein RR, Gibson SI (2002) ABA and sugar interactions regulating development: cross-talk or voices in a crowd? Curr Opin Plant Biol 5(1):26–32. https://doi.org/10.1016/s1369-5266(01)00225-4
León P (2003) Sugar and hormone connections. Trends Plant Sci 8(3):110–116. https://doi.org/10.1016/s1360-1385(03)00011-6
Cheng WH, Endo A, Zhou L et al (2002) A unique short-chain dehydrogenase/reductase in Arabidopsis glucose signaling and abscisic acid biosynthesis and functions. Plant Cell 14(11):2723–2743. https://doi.org/10.1105/tpc.006494
Zhang Z, Mao CY, Shi Z et al (2017) The amino acid metabolic and carbohydrate metabolic pathway play important roles during salt-stress response in tomato. Front Plant Sci 8:1231. https://doi.org/10.3389/fpls.2017.01231
Inskeep WP, Bloom PR (1985) Extinction coefficients of chlorophyll a and b in N, N-dimethylformamide and 80% acetone. Plant Physiol 77(2):483–485. https://doi.org/10.1104/pp.77.2.483
Zhang ZQ, Pang XQ, Duan XW et al (2005) Role of peroxidase in anthocyanin degradation in Litchi fruit pericarp. Food Chem 90(1–2):47–52
Alici E, Arabaci G (2016) Determination of SOD, POD, PPO and CAT enzyme activities in Rumex obtusifolius L. Annu Res Rev Biol 11(3):1–7. https://doi.org/10.9734/arrb/2016/29809
Mohammadkhani N, Heidari R (2008) Drought-induced accumulation of soluble sugars and proline in two maize varieties. World Appl Sci J 3(3):448–453
Kou XH, Chen XY, Mao CY et al (2019) Selection and mechanism exploration for salt-tolerant genes in tomato. J Hortic Sci Biotechnol 94(2):171–183. https://doi.org/10.1080/14620316.2018.1486739
Akram NA, Noreen S, Noreen T et al (2015) Exogenous application of trehalose alters growth, physiology and nutrient composition in radish (Raphanus sativus L.) plants under water-deficit conditions. Braz J Bot 38(3):431–439. https://doi.org/10.1007/s40415-015-0149-7
Moradi F, Ismail AM (2007) Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Ann Bot 99(6):1161–1173. https://doi.org/10.1093/aob/mcm052
Husen A, Iqbal M, Aref IM (2016) IAA-induced alteration in growth and photosynthesis of pea (Pisum sativum L.) plants grown under salt stress. J Environ Biol 37(3):421–429
Tavakoli M, Poustini K, Alizadeh H (2018) Proline accumulation and related genes in wheat leaves under salinity stress. J Agr Sci Tech-Iran 18(3):707–716
Huang ZR, Zhao L, Chen DD et al (2013) Salt stress encourages proline accumulation by regulating proline biosynthesis and degradation in Jerusalem artichoke plantlets. PLoS ONE 8(4):e62085
Miller GAD, Suzuki N, Ciftci-Yilmaz S et al (2010) Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell & Environ 33(4):453–467
Durand M, Porcheron B, Hennion N et al (2016) Water deficit enhances C export to the roots in Arabidopsis thaliana plants with contribution of sucrose transporters in both shoot and roots. Plant Physiol 170(3):1460–1479
Parihar P, Singh S, Singh R et al (2015) Effect of salinity stress on plants and its tolerance strategies: a review. Environ Sci Pollut Res 22(6):4056–4075
Liang WJ, Ma XL, Wan P et al (2018) Plant salt-tolerance mechanism: a review. Biochem Biophys Res Commun 495(1):286–291. https://doi.org/10.1016/j.bbrc.2017.11.043
Li HW, Zang BS, Deng XW et al (2011) Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta 234(5):1007–1018
Li NN, Qian WJ, Wang L et al (2017) Isolation and expression features of hexose kinase genes under various abiotic stresses in the tea plant (Camellia sinensis). J Plant Physiol 209:95–104
Sami F, Yusuf M, Faizan M et al (2016) Role of sugars under abiotic stress. Plant Physiol Biochem 109:54–61
Hummel I, Pantin F, Sulpice R et al (2010) Arabidopsis plants acclimate to water deficit at low cost through changes of carbon usage: an integrated perspective using growth, metabolite, enzyme, and gene expression analysis. Plant Physiol 154(1):357–372
Durand M, Mainson D, Porcheron B et al (2018) Carbon source-sink relationship in Arabidopsis thaliana: the role of sucrose transporters. Planta 247(3):587–611
Gong X, Liu ML, Zhang LJ et al (2013) Sucrose transporter gene AtSUC4 responds to drought stress by regulating the sucrose distribution and metabolism in Arabidopsis thaliana. Adv Mater Res 765–767:2971–2975
Kong XQ, Luo Z, Zhang YJ et al (2017) Soaking in H2O2 regulates ABA biosynthesis and GA catabolism in germinating cotton seeds under salt stress. Acta Physiol Plant 39:2. https://doi.org/10.1007/s11738-016-2299-z
Shu K, Qi Y, Chen F et al (2017) Salt stress represses soybean seed germination by negatively regulating GA biosynthesis while positively mediating ABA biosynthesis. Front Plant Sci 8:1372. https://doi.org/10.3389/fpls.2017.01372
Saeedipour S (2012) Is salinity tolerance of rice lines concerned to endogenous ABA content or to the cellular ability for ABA synthesis under stress? Afr J Biotechnol 11(49):10938–10943
Thompson AJ, Andrews J, Mulholland BJ et al (2007) Overproduction of abscisic acid in tomato increases transpiration efficiency and root hydraulic conductivity and influences leaf expansion. Plant Physiol 143(4):1905–1917
Acknowledgements
This study was supported by the National Natural Science Foundation of China (Nos. 31671899 and 31871848). We are very grateful to Dr. Daqi Fu (China Agricultural University) for providing the tomato seeds.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Yanchun Feng and Xiuyu Chen contributed equally to this work.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Feng, Y., Chen, X., He, Y. et al. Effects of Exogenous Trehalose on the Metabolism of Sugar and Abscisic Acid in Tomato Seedlings Under Salt Stress. Trans. Tianjin Univ. 25, 451–471 (2019). https://doi.org/10.1007/s12209-019-00214-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12209-019-00214-x