
Abstract
Temperature stress impacts both welfare and productivity of livestock. Global warming is expected to increase the impact, especially in tropical areas. We investigated the biological mechanisms regulated by temperature stress due to the circadian temperature cycle in temperature adapted and non-adapted chicken under tropical conditions. We studied transcriptome profiles of heart, breast muscle, and spleen tissues of Ethiopian lowland chicken adapted to high circadian temperatures and non-adapted Ethiopian highland chicken under lowland conditions at three points during the day: morning, noon, and evening. Functional annotations and network analyses of genes differentially expressed among the time points of the day indicate major differences in the reactions of the tissues to increasing and decreasing temperatures, and also the two chickens lines differ. However, epigenetic changes of chromatin methylation and histone (de)acetylation seemed to be central regulatory mechanisms in all tissues in both chicken lines. Finally, all tissues showed differentially expressed genes between morning and evening times indicating biological mechanisms that need to change during the night to reach morning levels again the next day.
Similar content being viewed by others


Introduction
Environmental changes, especially temperature changes, influence animal health, welfare, and livestock productivity. Temperatures above normal are felt as heat stress (HS) by all living organisms, and heat stress can be defined as the sum of all external forces acting on an animal that causes an increase in body temperature and evokes a physiological response (Fuquay 1981; Phillips and Piggins 1992, Dikmen and Hansen 2009). For example, it has been shown that the level of productivity and health of dairy cattle, goat, and sheep and layer and broiler chicken are reduced by heat stress, and chicken mortality is increased (Fuquay 1981; Bottje and Harrison 1985; Lu 1989; Young 1990; Yahav et al. 1995; Silanikove 1992; West 2003; Marai et al. 2007). Physiologically, this may relate to reduced feed intake, suppressed immune response, and changed gene expression profiles and animal metabolism (Harrison and Biellier 1969; Emery et al. 1984; Altan et al. 2003; Mashaly et al. 2004; Lin et al. 2006a; Srikanth et al. 2017).
In tropical environments, the daily temperature rise may induce heath stress to chicken. Local environmental conditions, such as altitude and natural conditions, e.g., forests, affect the level and the impact of the temperature rise during the day. Global warming may increase the local temperatures further increasing heat stress effects. This may especially affect future livestock productivity in sub-Saharan countries (Thornton et al. 2009).
The impact of heat on broiler and layer strains of chicken has been studied before (Zahoor et al. 2017). The impact of heat stress in both genotypes of poultry is very similar (Zahoor et al. 2016, 2017). Apart from decreased productivity levels, physiological and behavioral changes were noted affecting not only animal welfare but potentially also consumer health (Lara and Rostagno, 2013). Furthermore, acute heat stress-affected gene expression profiles in the testis of broiler chickens increased the number of apoptotic cells, suggestive for reduced fertility (Wang et al. 2015). Changed gene expression in muscle tissue of broiler chickens has also been reported, suggestive for reduced meat productivity (Li et al. 2011).
As with most environmental challenges, animals living for generations in hot climates adapt to the environmental conditions (Lin et al. 2006b; Renaudeau et al. 2012; Lara and Rostagno, 2013; Kim et al. 2017). Chickens can adapt to circadian heath cycles (Lin et al. 2006b; Renaudeau et al. 2012; Lara and Rostagno, 2013). Furthermore, increasing the temperature to 1–1.5 °C during days 8–21 of egg incubation induced an adaptive response to hot temperatures in later life (Moraes et al. 2003; Janke and Tzschentke, 2010; Loyau et al. 2014a, b, 2015, 2016), including changed genome activity, suggesting epigenomic mechanisms (Loh et al. 2004; Yossifoff et al. 2008).
Together, these data suggest that the transcriptomic changes mediating the response of the chicken to heat stress differ between adapted and non-adapted chickens. We here report on the differences in the transcriptomic profiles and the underlying regulated biological mechanisms in three vital tissues: heart, muscle (meat productivity), and spleen (immune) using a model system of chicken adapted to hot environments and non-adapted chickens. The objectives of these studies were to determine (1) the transcriptome profile differences between highland and lowland Ethiopian chicken before, during, and after hot daytime temperatures and (2) the transcriptome profile changes in highland and lowland Ethiopian chicken separately during the day.
Materials and methods
Animals and experimental design
The care and experimental use of Ethiopia chicken were reviewed and approved by the Institutional Animal Care and Use Committee at the International Livestock Research Institute (ILRI, Ethiopia) (IACUC No. 2016-216).
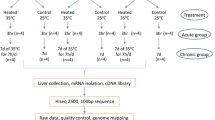
Ethiopian highland male chickens (N = 9) from Addis Ababa (altitude 2400 m above sea level, average high temperature 16 °C) were collected randomly and transferred to the Ethiopian lowland Awash village (altitude 950 m above sea level, average high temperature 37 °C), where the experiments were performed after a 20-h free-range period to relieve the animals from the transportation stress. Similarly, Ethiopian male lowland chicken from Awash village (Yangji hatchery, N = 9) were collected randomly. The Ethiopian endogenous chicken lines originated from the same route to Ethiopia but separated to different environmental conditions many generations ago. The lowland chickens were adapted to the circadian high temperature cycle, while the highland chickens were not.
All chickens were subjected to the lowland circadian temperature cycle at the same day. The chickens were slaughtered at the experimental location at three different time points: early in the morning at 6.00 AM (i.e., before the onset of increasing temperature; measured conditions 25.2 °C, 64% humidity), at noon, 12.00 PM (i.e., the hottest temperature; measured conditions 34.6 °C, 6% humidity), and in the evening at 6.00 PM (i.e., after the temperature decline; measured conditions 26.3 °C, 9% humidity). Three chickens of each group were sacrificed per time point. Heart, breast muscle, and spleen tissue were snap-frozen and stored at − 80 °C until experimentation. The tissues were transferred to the Republic of Korea for RNA-Seq analysis.
RNA-Seq analysis
RNA was isolated using TRIzol (Invitrogen, USA). Hundred milligrams of total RNA of each tissue were used to construct libraries for sequencing. The mRNA-Seq library was constructed with TruSeq mRNA library (Illumina Inc.) and sequenced with Illumina HiSeq2000 100PE (Illumina, Inc., San Diego, CA, USA) sequencing in accordance with the manufacturer’s instructions. To remove rRNA, bead-based depletion was used. Libraries were sequenced independently. Obtained sequence reads were aligned to the Gallus gallus reference genome (NCBI GenBank, ftp://ftp.ensembl.org/pub/release-89/fasta/gallus_gallus/dna/). Preprocessing and mapping were performed using the pipelines employing FastQC, trimmomatic, and Tophat2. The read mapping was conducted using the RNA-Seq analysis package default settings (mismatch cost 2, insertion cost 3, length fraction 0.8, similarity fraction 0.8, and maximum number of read 1) within the CLC software (Qiagen, Redwood city, CA, USA; https://www.qiagenbioinformatics.com/products/clc-genomics-workbench/). The number of mapped reads was normalized and measured in terms of reads per kilobase per million (FPKM) to determine transcript abundance using Cufflinks.
Bioinformatic analysis
This experiment investigates the reaction to circadian environmental temperatures in a hot climate zone using two groups of chicken differing for adaptation to high environmental temperatures. Therefore, the first analysis is the direct comparison between the two groups of chicken at each time point.
Each chicken type was investigated at three time points: morning, noon, and evening (see above). Thus, for each chicken type, three analyses were done comparing morning–noon (i.e., the reaction to increasing temperatures), noon–evening (i.e., the reaction to decreasing temperature), and morning–evening (i.e., to investigate whether recovery from hot temperature is complete or that further recovery is needed during night time).
For each analysis, functional annotation analysis and network analyses were done. Because the analysis of the results using the chicken data was unsatisfying, we converted the ENSEMBLE gene numbers to official gene symbols and used the human annotation data. We compared the results obtained with the human annotation always with the results obtained with the chicken annotation. However, here we presented the results obtained with the human annotation. First differentially expressed genes were selected. For each gene, the expression levels were compared between the two groups. These lists of differentially expressed genes were used for functional annotation enrichment analysis using the DAVID software (Huang et al. 2009a, b; https://david.ncifcrf.gov/). Network analyses were done using the online software tools STRING (Szklarczyk et al. 2015, 2017; https://string-db.org/) and STITCH (Kuhn et al. 2014; Szklarczyk et al. 2016; http://stitch.embl.de/). The protein network was investigated with STRING, while the importance of specific metabolites was investigated with STITCH. The higher order pathway organization was investigated using the ClueGo app of Cytoscape (Shannon et al. 2003; Bindea et al. 2009). Groups of pathways were studied using KEGG and BioCarta pathway databases (Kanehisa and Goto, 2000; Kanehisa et al. 2014; http://www.genome.jp/kegg/; Nishimura, 2004; https://cgap.nci.nih.gov/Pathways/BioCarta_Pathways). The cutoff value for a group was arbitrarily set to 3. Several datasets were too large to analyze for the software due to large numbers of differentially expressed genes. To solve this, the software was set to maximum specificity.
Results
Differentially expressed genes and related functional annotation enrichment
Supplementary files 1, 2, and 3 provide all RNA-Seq data of all genes after normalization for respectively heart, muscle, and spleen tissues. These were the data used for all analyses. Table 1 showed that the numbers of differentially expressed genes differ among chicken groups and tissues. The difference between highland and lowland chickens seemed largest in muscle tissue. Lowland chickens showed the largest number of differentially expressed genes in heart tissue while highland chicken showed largest number of differentially expressed genes in spleen and muscle tissue. This may indicate differential biological regulatory mechanisms operating in the two chicken groups. Furthermore, all tissues showed reaction to increasing temperatures (i.e., morning–noon) and to decreasing temperatures (i.e., noon–evening). The comparison between gene expression in the morning and the evening indicates that at the end of the day no (full) reversal of gene expression has taken place, suggesting further changes in gene expression patterns during the night. Especially in lowland chicken heart tissue, major differences between morning and evening gene expression profiles were observed. In lowland chicken heart tissue, the reaction during increasing temperatures seemed to be much smaller than during decreasing temperatures. The highland versus lowland chicken comparison suggested a similar mechanism in muscle tissue, which already starts at noon.
Table 2 showed the number of significantly enriched functional annotations reported by DAVID. The number of enriched functional annotations using the chicken genome annotation is poor as compared to the human genome annotation data. Therefore, we used the functional annotation of the human genome, although we are aware of potential differences between mammals and birds.
Heart tissue of lowland chicken showed more functional annotation enrichments in lowland chicken than in highland chicken. Furthermore, lowland chicken showed the highest number of functional annotation enrichments during decreasing temperatures, which was not observed in highland chicken. Spleen tissue showed the opposite pattern with highest number of functional annotations enriched in highland chicken and largest number of enriched functional annotations during the period of increasing daytime temperatures. No differences between the chicken groups were found in muscle tissue. Again, all data indicate a major difference between morning and evening.
Functional annotations
In this paragraph, we focus on the biological processes underlying the data of Table 2. A summary of the functional annotations can be found in the Supplementary file 4. The data in the file focus on biological and cellular molecular processes and exclude the long lists of protein domains and interactions reported by DAVID but include the cluster analysis of DAVID. Functional annotations describing the same biological functionality were taken together, thus reducing the number of annotations without losing biological information. We describe the data per tissue and focus mainly on general mechanisms.
Heart tissue
Functional analysis of the differentially expressed genes between highland and lowland chicken indicates a role for mitochondrial processes, mainly related to oxidative energy metabolism, specifically at noon. The annotation was found also in lowland chicken in the afternoon period. In highland chicken during the morning period and in lowland chicken during the afternoon period and the morning–evening comparison, the functional annotation relating to cellular communication pathways (collagen/extracellular matrix/cell-cell adhesion and communication) is found. RNA metabolism, especially splicing, is found in both chicken groups. The chicken groups have differential expression at noon and in the evening. While highland chicken show differential expression during the day, lowland chicken indicate especially a difference between the morning and evening analyses. This indicates that in highland chicken, the expression is regulated during the day and returns to morning levels, but in lowland chicken, the expression between morning and evening remains different. Only in highland chicken during the morning to noon period that differential expression of ribosomal proteins is found. Finally, at several places, heart/muscle annotations are indicated suggesting rebuilding or restructuring of the heart tissue, which may relate to functional adaptations of the heart. Histone (de)acetylation and chromatin methylation were found in both chicken lines.
Muscle tissue
Analysis of the differentially expressed genes between highland and lowland chicken indicate splicing as an important mechanism in both chicken groups, and in both groups, the expression in the evening did not return to the expression levels in the morning. The highland and lowland chicken groups differ in expression of the proteasome complex, especially during the afternoon period. This difference related mainly to the lowland chickens, which also indicate that expression levels also differ between morning and evening samples. The highland and lowland chicken groups differ also in expression for the cellular communication pathways (collagen/extracellular matrix/cell-cell adhesion and communication), especially during the afternoon period. These effects were mainly found in the highland chickens and not in the lowland chickens. The two chicken group also differed for the expression of mitochondrial genes, which seemed to be regulated only in the highland chicken, which also indicated that evening expression levels differ from morning expression levels. The two chicken groups also differed for acetylation, especially during the afternoon period. Differential expression was observed throughout the day in both chicken groups, indicating that expression remained different between evening and morning expression levels. Finally, at several places, muscle annotations were indicated suggesting rebuilding or restructuring of the muscle tissue, which may relate to functional adaptations of the muscle tissue.
Spleen tissue
Analysis of the differentially expressed genes between highland and lowland chicken indicates functional annotation enrichment of mitochondrial genes at noon. The expression profiles of the two chicken groups differ: in highland chicken, differentially expressed mitochondrial genes were found during the morning period, while in lowland chicken, functional annotation enrichment of mitochondrial genes was found during the afternoon period. Only in highland chicken the comparison of the expression levels of genes for the evening to morning showed differentially expressed genes. It should be noted that here especially mitochondrial organization was involved, while no oxidative phosphorylation mechanism annotations were reported. Functional annotation enrichment of cellular communication-related pathways (collagen/extracellular matrix/cell-cell adhesion and communication) was found between highland and lowland chickens, especially in the evening. Some indications for differential regulation of these processes were found in highland chicken but not in lowland chicken. These data are supported by regulation of post-translational modification of proteins (Golgi/endoplasmic reticulum/protein transport) data. The two chicken groups differed especially in the morning, while specific regulation was found throughout the day in both chicken groups. Especially in lowland chicken, regulation of alternative splicing was found throughout the day. The two chicken groups differed in the expression of acetylation-related genes at noon. However, both groups showed differential expression of acetylation-related genes throughout the day, with some indication of histone-specific acetylation/deacetylation. Finally, enrichment of cell division was observed in both chicken groups in the morning but not in the afternoon.
Network analyses
Three different network analyses were done: a protein network analysis, a protein network analysis including metabolites, and an integration of pathway analyses. Supplementary file 5 provides an overview of the results in the three tissues. All network analysis figures are provided in Supplementary file 6. For the protein networks, we focused on the hub proteins defined as highly connected proteins as compared with other proteins in the network. Hub proteins were analyzed for functional annotation, and the functional annotations of the hub proteins are displayed in Table 3. Furthermore, we observed in several networks specific nuclei, defined as groups of proteins connected with each other but with less connections to other proteins. The proteins of the nuclei were analyzed for functional annotation—it proved to be that the proteins within a nucleus had similar functional annotation. Several of the network analyses were hampered by the creation of very dense networks, too-large-to-analyze. It should be noted that from these networks, data may be missing or incomplete.
Table 3 shows a summary of the functional annotation enrichments of the protein hubs and nuclei of the networks of the three tissues at three different times of the day. The table shows an extraction of the data that can be found in detail in the Supplementary files 2 and 3. The table shows that several general functional mechanisms are shared by the tissues: energy metabolism, (de)acetylation, mitosis/apoptosis, immune functions, circadian rhythm, (molecular) chaperone, stress, and (alternative) splicing. Double annotations suggest that stress and splicing and stress (molecular), chaperone, and mitosis/apoptosis are related to each other. Furthermore, several metabolites were found to be related to functional mechanisms of differentially expressed genes. Among these, calcium and dopamine are shared among the tissues.
Groups of pathways related to heat stress changes throughout the day
Table 4 shows the results of the Cytoscape-ClueGo analysis of groups of (physiological) pathways harboring differently expressed genes in three tissues: heart (A), (breast) muscle (B), and spleen (C). In heart tissue, the main remarkable result is that lowland chicken seems to respond more than the highland chicken. The lowland chicken shows differential expression in positive regulation of several metabolic pathways. Despite this, there is a long list of groups of pathways differing between morning and evening, indicating that the heat-related changes were not reversed yet at the end of the day during the afternoon. Among them, negative regulation of histone H3-K4 methylation indicates potential increased gene expression levels. In the afternoon, a relation with the gut is suggested via the vagus nerve. The circadian rhythm is changed in highland chicken during the day and still changed at the end of the day. Major differences between the highland chicken and the lowland chicken also include cardiac and skeletal muscle developmental effects, fat cell differentiation and lipid storage, and cell division. There seems to be a relation with immune-related processes and auditory signal responses (especially in the highland chicken).
Muscle tissue shows several main themes in both chicken lines, including (1) histone modifications, especially during the morning; (2) blood vessel development, in highland chicken mainly during the morning and in lowland chicken mainly during the afternoon; (3) cell division, at the end of the day the situation still differs from the beginning of the day; (4) oxidative stress, in highland chicken especially during the morning; (5) apoptosis throughout the day; (6) response to stress, throughout the day, and in both chicken lines the evening situation still differs from the morning situation; (7) nerve activity, in the lowland chicken mainly during the morning, but the situation at the end of the day still differs from the beginning of the day; and (8) immune response changes. The two chicken lines also differed for muscle differentiation processes and muscle activity. Especially, lowland chicken had a difference in photoperiodism in the morning.
Spleen tissue showed very few differences between the two chicken lines, and in the evening, and they seemed to be similar. Some differences may include regulation of cell division, learning, protein localization at noon, and ear development and functioning. Despite of this, the analysis of both chicken lines individually indicated more differences. Highland chickens showed a wide array of biological processes during the day, with the main focus of regulated processes during the morning. This includes the biological processes leading to apoptosis, histone H3 acetylation, inner ear development, and cell-cell contacts and cell cycle checkpoints. At the end of the day, the processes of apoptosis and cell division were still different from the morning expression levels. Other enriched biological mechanisms were energy metabolism and angiogenesis—which may be related to water loss, blood pressure, cardiovascular disease, and IL4 biosynthesis groups of pathways. Contrarily, the lowland chicken line showed regulation of the proinflammatory IL17 production specifically during the morning. Lowland chickens also showed regulation of energy metabolism more widely via fatty acid, cholesterol, and energy metabolism. In general, they showed regulation of the groups of pathways of circadian rhythm and cellular response to heat and stress in general and oxidative stress specifically—which is still regulated in the evening. These all may be related to DNA hypermethylation.
Heat stress proteins related to heat stress—differences between chickens and tissues during the day
We investigated the differential expression of the heat shock proteins (HSPs) because these are especially related to the physiological reaction to these specific environmental conditions. We observed differences among the tissues over the day. In heart tissue, we found that the two chickens differed in expression of HSPs especially during the afternoon. Functionally, these HSPs are involved in mitochondrial electron transport and others function as molecular chaperones in the nucleus, regulating cell cycle control and signal transduction. The lowland and highland chickens differ in that the mitochondrial HSP is regulated in lowland but not in highland chicken. On the contrary, in highland chicken, a HSP transcription factor and cell cycle control and signal transduction regulating HSP were regulated, which were not regulated in lowland chicken. However, the chickens share regulation of HSP functioning as a molecular chaperone for correct folding of proteins and controlling protein transport in the ER. Calcium availability is important for this.
The regulated HSP gene profiles in the muscle tissue were different. First of all, the two chickens showed different HSP regulation throughout the day, especially for HSPs functioning as transcription regulators, cell cycle control, and signal transduction and especially for HSPs recruited to the nucleus under heat stress conditions. Furthermore, in the afternoon, the chickens differ for regulation of glucocorticoid and mitochondrial electron transport signaling. At the end of the day, a muscle-specific HSP is differently expressed involved in regulation of actin polymerization. In both chickens, no HSPs regulated during the afternoon. Finally, it is remarkable that there were many more HSPs regulated in highland chickens than in lowland chickens. The focus in lowland chicken is on regulation of heat shock transcriptions factors during the morning and at the end of the day. In highland chickens, this was also found during the morning, but throughout the day, environmental stress-regulated HSPs were found functioning as chaperones and regulating the cellular response to heat and protecting from apoptosis. The muscle-specific HSP was found regulated at the end of the day.
Spleen tissue showed only differences between the two chickens in the afternoon, where the two chickens differed for HSPs regulating mitochondrial electron transport and cellular molecular chaperones described especially in the cellular response to heat stress. However, when investigating the two chickens individually, they showed different profiles. While lowland chickens showed regulated HSPs throughout the day, highland chickens did not have regulated HSPs in the afternoon. Lowland chickens regulated HSPs functioning for glucocorticoid signaling and molecular chaperones especially protecting against aggregated proteins. Highland chickens especially regulated HSPs during the morning. These HSPs especially regulated functions related to signaling to the innate immune system and controlling correct toll-like receptor folding in the endoplasmic reticulum. A relation with the melanosome was also suggested.
Visualization of circadian expression profiles in three tissues of highland and lowland chicken
Muscle cell integrity and functioning
The expression profiles of several proteins suggest that the integrity of the muscle tissue may be affected during circadian heat. The expression profiles of the serpin peptidase inhibitor (Serpine, Clade E) gene involved in cellular adhesion and extracellular remodeling, the collagen type V alpha 2 chain and collagen type VI alpha 3 chain (Col5A2 and Col6A3, respectively) gene involved in signaling and focal adhesion, and the dermatopontin (DPT) gene involved in cell-matrix interactions and matrix assembly all follow the same expression pattern showing that the expression in the muscle tissue is decreased during the day in highland chickens but not in lowland chickens. The consequence may be that in highland chickens the extracellular matrix of muscle cells is less well organized, and as a consequence the muscle functioning may be less. We have shown similar effects on the muscle tissue during overtraining of horses (te Pas et al. 2013). Overtraining may also related to body heat. Figure 1a shows the expression pattern of the Col5A2 as an example of this expression profile. Please note that the extracellular matrix protein 2 (ECM2) gene shows the same expression protein but also in the heart tissue of highland chickens.




Visualization of circadian expression profiles in three tissues of highland and lowland chicken. a Collagen type V alpha 2 chain (Col5A2). b Heat shock protein B3 (HSPB3). c RNA binding motif protein 15 (RBM15). d Pyruvate dehydrogenase kinase, isoenzyme 4 (PDK4). e Calcium binding protein 1 (CABP1). f DnaJ heat shock protein family (Hsp40) member C12 (DNAJC12). g ETS2 repressor factor (ERF). h Inhibitor of growth family member 2 (ING2). i Nuclear receptor subfamily 4 group A member 3 (NR4A3). j Biliverdin reductase A (BLVRA)
This effect may be strengthened via heat shock protein expression. Figure 1b shows the expression pattern of the muscle-specific heat shock protein HSPB3. In highland chickens, the expression profile is the same as described above, while the opposite expression profile is observed in lowland chicken, suggesting protection from heat stress in lowland chickens.
The RNA minding motif protein 15 (RBM15) gene expression profile shows a sharp increase at noon in highland chickens. Since the gene is involved in repressing several signaling pathways, this further strengthens the hypothesis of loss of muscle tissue function (Fig. 1c).
Energy expenditure
Heat stress may affect the energy metabolism because animals may move less and eat less during heat stress. The mitochondrial pyruvate dehydrogenase kinase, isoenzyme 4 (PDK4) gene expression profile is shown in Fig. 1d. The profile has shown that the expression in all tissues is lower at noon. The expression levels in the morning and the evening are approximately the same suggesting that full recovery of the effect during increasing temperature is taking place in the afternoon when temperature is reduced. The protein is also important for maintaining normal blood pH and in preventing the accumulation of ketone bodies under starvation, which may relate to less feeding at noon.
Metabolism
The calcium binding protein 1 (CABP1) gene is involved in calcium-mediated cellular signal transduction regulating numerous processes. Its expression profile (Fig. 1e) shows a high peak at noon in highland chicken while lowland chickens show no regulation in muscle tissue. The DnaJ heat shock protein family (Hsp40) member C12 (DNAJC12) gene shows downregulation in the heart tissue of highland chicken but not in lowland chicken (Fig. 1f). This heat shock protein is involved in complex assembly, protein folding, and export. The profile suggests that the protein metabolism of the heart of highland chicken is affected. Apoptosis may be involved in all tissues and in both chickens. Figure 1g shows the expression profile of the ETS2 repressor factor (ERF) gene suggesting increased expression in both chickens and all three tissues.
The inhibitor of growth family member 2 (ING2) gene is involved in modulation of the activity of histone acetyltransferase and histone deacetylase complexes regulating gene expression and function in DNA repair and apoptosis (Fig. 1h). The expression profile shows a major downregulation of the expression especially in the heart tissue of lowland chickens. Regulation of the expression via this mechanism may be an adaptation to heat stress in lowland chickens. The nuclear receptor subfamily 4 group A member 3 (NR4A3) gene (Fig. 1i) has a role in the regulation of proliferation, survival, and differentiation of many different cell types and also in metabolism and inflammation. The expression profile shows a sharp decreased expression in heart and muscle tissues of highland chickens, while the lowland chickens show the opposite expression profiles in both tissues.
The biliverdin reductase A (BLVRA) gene is an example of a gene regulated in spleen tissue (Fig. 1j). The expression is downregulated during the day, in highland chicken already at noon and in lowland chicken during the afternoon. The gene is involved in heme catabolism and may therefore relate to blood oxygen transport capacity.
Discussion
We investigated the biological mechanisms underlying the reaction to daytime heat stress in chicken. Regular diurnal changes in physical activity will also affect gene expression profile changes throughout the day. Therefore, we used two chicken lines differing for heat adaptation. We used a lowland chicken line used to high noon temperatures and a highland chicken line not used to this. Both chicken lines were exposed to lowland temperature conditions at the same day. The highland chickens were transported to the lowland location and rested for 20 h before the onset of the experiment. This recovery time is vital because if the recovery time is too short the effect of transportation is part of the measured response, and if the recovery time is too long the first signs of adaptation may be measured. But what is the best recovery time? Part of the answer may be found in the lairage time after transportation before slaughtering an animal. This is usually 3 h. In broilers, it is called “long-term recovery,” and it is effective in lowering plasma corticosterone levels (Zhang et al. 2009). Therefore, it may be assumed that this time effectively reduces transportation stress. However, this work is not about slaughtering the chickens but performing an experiment measuring circadian stress in relation to the (absence of) adaptation to high daytime temperatures. In our choice for 20 h resting time, we are sure that transportation stress is effectively reduced while the animals did not have to time to build up an adaptation response.
We expected differences between the chicken lines in their reactions to lowland daytime temperature variations. Therefore, we analyzed both chicken lines early in the morning, at noon, and late in the evening, i.e., before the daily rise in temperature, at the hottest moment, and after the decline in temperature. We measured the gene expression levels of three tissues: heart, muscle, and spleen. The heart is an organ that has been shown to react to heat stress and is involved in adaptation to environmental heat (Lin et al. 2006a, b). The breast muscle in chicken is the most valuable piece of meat and therefore important for productivity, and the spleen is important for immune functionality. Our results also indicated interactions among these organs showing the importance of investigating several tissues similarly to highlight the biological mechanisms regulated body wide. We compared the gene expression levels among day time points within each chicken line and per time point between the two chicken lines.
We showed differently expressed genes at each time point in each tissue of both chicken lines. The number of differently expressed genes and the (number of) functional annotation enrichments vary among chickens lines and time points. Not surprisingly, there is a high similarity in the patterns of the number of differently expressed genes and the functional annotation enrichments of the biological mechanisms at a higher regulatory level. In highland chicken, the number of differently expressed genes and functional annotations is spleen > muscle > heart, while lowland chicken showed heart > muscle > spleen. When comparing the differently expressed genes between the two chicken lines, we found muscle > heart > spleen. This shows directly that the two chicken lines differ in reaction to heat stress and that this difference is tissue-specific.
Next, we investigated the effect of time of the day related to circadian temperature variation. Here, we found major differences between the highland and the lowland chicken. In the heart tissue of highland chicken, the largest number of regulated biological mechanisms is in the morning, less in the afternoon, and only a low number of biological mechanisms remaining in the evening. In lowland chicken, this is the reverse. This suggests that the heart tissue of highland chicken reacts early to increased temperatures, while the heart tissue in lowland chicken reacts later, especially during declining temperatures in the afternoon. It cannot be excluded, however, that this was still a reaction to increasing temperatures in the morning but late. This correlates well with the observation that in the evening heart tissue was very different from the morning. Because we suggest that the next day the situation in the morning should have been restored, these data indicate that during the night, lowland chicken will need to regulate many biological processes. Since lowland chicken are used to the high noon temperatures, this may be an adaptation mechanism in heart tissue.
In muscle tissue, highland chicken showed similar levels of regulation throughout the day, while lowland chicken showed more regulation in the morning, suggesting that here lowland chicken reacts early while highland chicken react both early and late. However, since in the morning highland chicken and lowland chicken were highly similar, it is supposed that both chicken lines have similar biological-regulated mechanisms. During the afternoon, both chicken lines started to be different and a relatively high number of biological mechanisms differ between evening and morning situation in both chicken lines, suggesting that both chicken lines need to have repair mechanisms in muscle tissue during the night. Muscle tissue has a high volume, especially in broiler-type chicken, and this may require more time to respond to environmental heat and reverse the reaction in the afternoon.
Spleen tissue is remarkably different from the other two tissues, especially in highland chicken. This indicated that regulation of immune processes differed between the two chicken lines. The number of differentially regulated functional annotation enrichments is much higher in highland chicken than in lowland chicken in the morning. Thus, major immune process changes were taking place during rising temperatures in the chicken that were not used to high noon temperatures, while this was not the case in adapted chicken. In highland chickens, the number of genes showing differential expression between morning and evening time points was low indicating that most processes take place during daytime.
This is in agreement with the high number of functional annotation enrichments differing between evening and morning in highland chicken. Surprisingly, the number of functional annotation enrichments differing between highland and lowland chicken was much lower. This suggests that many of the changed functional annotations in highland chicken belong to the same biological processes.
In conclusion, we showed major differences in the regulation of biological functional annotations related to increasing and decreasing temperatures during the day. These differences related to the tissue under investigation and the adaptation of chicken.
Biological mechanisms regulated in relation with circadian temperature changes
To investigate the biological processes underlying all these changes, we used functional annotation enrichment software and network analyses. For the latter, we used software to make protein networks and we focused on the hub genes and metabolites, and we studied groups of pathways. The added value of studying groups of pathways rather than single pathways is that it brings together more differently expressed genes and pathways. However, because of the sizes of several datasets, some networks were difficult to produce leading to loss of data for groups of pathways. However, data for the most important larger groups will remain. Data from these analyses were both confirming each other and were additive at other places.
Before discussing the regulated biological mechanisms per tissue throughout the day, we discuss a general theme found in our tissues in both chicken lines: epigenetics. Chromatin remodeling through both DNA methylation and histone (de)acetylation was reported. Especially transcriptional silencing was noticed, generally related to methylation and deacetylation (Malecová and Morris 2010; Weinberg and Morris 2016), although negative regulation of methylation leading to increased gene expression was also found. This can be a general biological process regulating all other biological mechanisms though transcription regulation. Epigenetic processes are of utmost importance for regulating genome-wide transcription activity (Orozco et al. 2015; Karsli-Ceppioglu et al. 2017). This suggest that the impact of temperature increase is such that genome-wide activity changes were induced, explaining the size of several of the datasets. Indeed, DNA methylation profile changes have been linked to response to stress in general (Unternaehrer et al. 2012) and to constant heat stress in particular (Hao et al. 2016), here we link it to circadian heat stress. Epigenomics is also a short-time regulator and much faster than mutation and selection (Berger 2007). Epigenetic regulation of the circadian clock has been reported (Sahar and Sassone-Corsi 2013), which makes it likely that also circadian-regulated processes are regulated via epigenetic mechanisms. Our results also indicate that the lowland chickens did not fully genetically adapt to the temperature conditions, although this epigenetic mechanism was found more in highland chickens than in lowland chickens, and epigenetic modifications may be inherited (Feeney et al. 2014; Orozco et al. 2015).
Finally, proteins especially regulated during environmental heat are the heat shock proteins. They perform functions dedicated to cellular survival. Indeed, we found HSPs involved in regulation of the energy metabolism, molecular chaperones ensuring correct protein folding for proper protein functioning, and muscle- and immune-specific functions. The tissues differ in these functions according to their different functions, i.e., immune functions were regulated in the spleen and the muscle tissue expressed the muscle-specific HSP required for proper actin polymerization. Especially the muscle tissue showed more HSP regulation in highland chicken as compared with lowland chicken suggesting that muscle tissue is especially vulnerable to heat stress, more than the other tissues under investigation. This is in agreement with our other muscle-specific data (see the Discussion below).
Heart tissue
Highland chicken heart tissue showed major regulation of protein synthesis in the morning that remains different until the evening. This may have affected the other reported processes including cell-cell contacts, circadian rhythm, apoptosis and mitosis, and cardiac and muscle developmental processes. This showed that heart tissue is under major remodeling during the day. The molecular chaperones and angiogenesis may add to this. Contrarily, lowland chicken mainly showed regulation of metabolic pathways: fatty acid synthesis and beta-oxidation and energy metabolism (mitochondrion). Cell-cell contacts and other cellular annotations were found only in the evening. This again shows that the heart tissue in lowland chicken differs between morning and evening. In both chicken lines, angiogenesis, circadian rhythm, and stress response mechanisms were observed indicating that daytime temperature is stressful for chicken hearts, regardless whether the conditions of the animals were adapted to changes in temperatures during the circadian cycle.
Muscle tissue
The data indicate both high similarity in biological processes between the two chicken lines and major differences of numbers of functional annotations between the two lines. This can be explained by the observation that similar processes were taking place in both chicken lines at different moments during the day. For this, we refer to the discussion above. Major general mechanisms include RNA (alternative) splicing protein synthesis and proteolysis, angiogenesis, nerve activity, apoptosis, and response to stress. This clearly showed that muscle tissue heavily reacts to environmental temperature. This has been shown for muscle tissue in several chicken lines before (Lara and Rostagno 2013; Zahoor et al. 2016, 2017). Especially in broiler chickens, muscle tissue is a heavily selected tissue. Moreover, the size and high metabolic activity of muscle tissue may lead to higher vulnerability to heat. More blood vessels in muscle tissue can support the loss of metabolically produced heat in the muscle tissue. This will also support nerve activity and tissue apoptosis. Together, it may improve the ability of the muscle tissue to react to heat stress properly. Consequently, there is a relation with immune processes.
Spleen tissue
As we have seen, both heart and muscle tissues have regulated immune-related biological mechanisms. Therefore, regulation of expression profiles of immune-related organs like the spleen was expected. Similarly, spleen functional annotations indicate cardiac-, muscle-, brain-, and ear-related regulated biological function. This indicated that the immune system has a widespread function, and the organs in an animal are connected as a functional unity (Martinez et al. 2015; Meng et al. 2016; Tidball 2017). This has been shown before, e.g., Veldman et al. (1984) reported a relation between spleen and ear disorders, and also in the traditional Chinese medicine spleen is seen as a regulator of muscle tissue and heart health (http://www.itmonline.org/5organs/spleen.htm).
Especially, the highland chicken showed a large array of different biological pathways in the morning. This was expected since the functional annotation indicated such a high number of regulated biological functions. However, when they were grouped, it appears that only a limited number of biological functions accounted for these annotations. A major portion relates to cellular division suggesting that the immune system is affected by the environmental temperature. Other major themes were protein metabolism and transport and molecular chaperones to maintain cellular integrity throughout the day. Several of these were also reported in cattle blood transcriptome analysis (Srikanth et al. 2017). Thus, cellular activity is regulated, and therefore immune capacity may be affected. A major difference between highland and lowland chickens is that highland chickens regulate IL4 biosynthesis, which regulates T helper 1 and 2, and B cell-cell activity in the immune response (Brown and Hural 1997), while lowland chickens regulate IL17, a T helper 2 cell activity and proinflammatory process regulator (Jin and Dong 2013). Although the consequence for immune capacity of the chickens is unknown at present, it may be expected that the animals have different capacity towards different pathogenic groups under heat stress conditions. In lowland chicken, energy metabolism and related metabolic pathways (fatty acid, cholesterol) is more regulated than in highland chicken, and the integration between energy metabolism and general metabolism is regulated. This suggests that the function of the immune cells may be affected. Angiogenesis and apoptosis appear general functions regulated in both chicken lines. These may be opposing biological functions. Angiogenesis will improve the functionality of the spleen, while apoptosis may negatively affect its functionality.
Acclimation to hot environment
In a world affected by global warming, environmental heat stress is a real threat to future livestock health and productivity. In this study, we showed that organ health, remodeling, and (metabolic) function may be affected but that the effects differ for each organ and each chicken line. Adaptation of animals to environmental heat may be a slow process and specific for animals with a different adaptation background.
References
Altan Ö, Pabuçcuoğlu A, Altan A, Konyalioğlu S, Bayraktar H (2003) Effect of heat stress on oxidative stress, lipid peroxidation and some stress parameters in broilers. Br Poult Sci 44:545–550
Berger SL (2007) The complex language of chromatin regulation during transcription. Nature 447:407–412
Bindea G, Mlecnik B, Hackl H, Charoentong P, Tosolini M, Kirilovsky A, Fridman WH, Pagès F, Trajanoski Z, Galon J (2009) ClueGO: a Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 25:1091–1093. https://doi.org/10.1093/bioinformatics/btp101
Bottje W, Harrison P (1985) Effect of carbonated water on growth performance of cockerels subjected to constant and cyclic heat stress temperatures. Poult Sci 64:1285–1292
Brown MA, Hural J (1997) Functions of IL-4 and control of its expression. Crit Rev Immunol 17:1–32
Dikmen S, Hansen PJ (2009) Is the temperature-humidity index the best indicator of heat stress in lactating dairy cows in a subtropical environment? J Dairy Sci 92:109–116
Emery DA, Vohra P, Ernst R, Morrison S (1984) The effect of cyclic and constant ambient temperatures on feed consumption, egg production, egg weight, and shell thickness of hens. Poult Sci 63:2027–2035
Feeney A, Nilsson E, Skinner MK (2014) Epigenetics and transgenerational inheritance in domesticated farm animals. J Anim Sci Biotechnol 5:48. https://doi.org/10.1186/2049-1891-5-48
Fuquay J (1981) Heat stress as it affects animal production. J Anim Sci 52:164–174
Hao Y, Cui Y, Gu X (2016) Genome-wide DNA methylation profiles changes associated with constant heat stress in pigs as measured by bisulfite sequencing. Sci Rep 6:27507. https://doi.org/10.1038/srep27507
Harrison P, Biellier H (1969) Physiological response of domestic fowl to abrupt changes of ambient air temperature. Poult Sci 48:1034–1045
Huang DW, Sherman BT, Lempicki RA (2009a) Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc 4:44–57
Huang DW, Sherman BT, Lempicki RA (2009b) Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res 37:1–13
Janke O, Tzschentke B (2010) Long-lasting effect of changes in incubation temperature on heat stress induced neuronal hypothalamic c-Fos expression in chickens. Open Ornithol 3:150–155
Jin W, Dong C (2013) IL-17 cytokines in immunity and inflammation. Emerg Microbes Infect 2(9):e60. https://doi.org/10.1038/emi.2013.58
Kanehisa M, Goto S (2000) KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res 28:27–30
Kanehisa M, Goto S, Sato Y, Kawashima M, Furumichi M, Tanabe M (2014) Data, information, knowledge and principle: back to metabolism in KEGG. Nucleic Acids Res 42:D199–D205
Karsli-Ceppioglu S, Dagdemir A, Judes G, Lebert A, Penault-Llorca F, Bignon Y-J, Bernard-Gallon D (2017) The epigenetic landscape of promoter genome-wide analysis in breast cancer. Sci Rep 7(6597):6597. https://doi.org/10.1038/s41598-017-06790-z
Kim J-M, Lim K-S, Byun M, Lee K-T, Yang Y-R, Park M, Lim D, Chai H-H, Bang H-T, Hwangbo J, Choi Y-H, Cho Y-M, Park J-E (2017) Identification of the acclimation genes in transcriptomic responses to heat stress of White Pekin duck. Cell Stress Chaperon 22:787–797. https://doi.org/10.1007/s12192-017-0809-6
Kotak S, Larkindale J, Lee U, von Koskull-Döring P, Vierling E, Scharf K-D (2007) Complexity of the heat stress response in plants. Curr Opin Plant Biol 10:310–316
Kuhn M, Szklarczyk D, Pletscher-Frankild S, Blicher TH, von Mering C, Jensen LJ, Bork P (2014) STITCH 4: integration of protein-chemical interactions with user data. Nucleic Acid Res 42:D401–D407
Lara LJ, Rostagno MH (2013) Impact of heat stress on poultry production. Animals 3:356–369. https://doi.org/10.3390/ani3020356
Li C, Wang X, Wang G, Li N, Wu C (2011) Expression analysis of global gene response to chronic heat exposure in broiler chickens (Gallus gallus) reveals new reactive genes. Poultry Sci 90:1028–1036. https://doi.org/10.3382/ps.2010-01144
Lin H, Decuypere E, Buyse J (2006a) Acute heat stress induces oxidative stress in broiler chickens. Comp Biochem Phys A 144:11–17
Lin H, Jiao H, Buyse J, Decuypere E (2006b) Strategies for preventing heat stress in poultry. Worlds Poult Sci 62:71–86. https://doi.org/10.1079/WPS200585
Loh B, Maier I, Winar A, Janke O, Tzschentke B (2004) Prenatal development of epigenetic adaptation processes in poultry: changes in metabolic and neuronal thermoregulatory mechanisms. Avian Poult Biol Rev 15:119–128
Loyau T, Métayer-Coustard S, Berri C, Crochet S, Cailleau-Audouin E, Sannier M, Chartrin P, Praud C, Hennequet-Antier C, Rideau N, Couroussé N, Mignon-Grasteau S, Everaert N, Duclos MJ, Yahav S, Tesseraud S, Collin A (2014a) Thermal manipulation during embryogenesis has long-term effects on muscle and liver metabolism in fast-growing chickens. PLoS One 9(9):e105339. https://doi.org/10.1371/journal.pone.0105339
Loyau T, Collin A, Yenisey Ç, Crochet S, Siegel PB, Akşit M, Yalçin S (2014b) Exposure of embryos to cyclically cold incubation temperatures durably affects energy metabolism and anti-oxidant pathways in broiler chickens. Poult Sci 93:1–9. https://doi.org/10.3382/ps.2014-03881
Loyau T, Bedrani L, Berri C, Métayer-Coustard S, Praud C, Coustham V, Mignon-Grasteau S, Duclos MJ, Tesseraud S, Rideau N, Hennequet-Antier C, Everaert N, Yahav S, Collin A (2015) Cyclic variations in incubation conditions induce adaptive responses to later heat exposure in chickens: a review. Animal 9:76–85
Loyau T, Hennequet-Antier C, Coustham V, Berri C, Leduc M, Crochet S, Sannier M, Duclos MJ, Mignon-Grasteau S, Tesseraud S, Brionne A, Métayer-Coustard S, Moroldo M, Lecardonnel J, Martin P, Lagarrigue S, Yahav S, Collin A (2016) Thermal manipulation of the chicken embryo triggers differential gene expression in response to a later heat challenge. BMC Genomics 17:329. https://doi.org/10.1186/s12864-016-2661-y
Lu C (1989) Effects of heat stress on goat production. Small Rumin Res 2:151–162
Malecová B, Morris KV (2010) Transcriptional gene silencing mediated by non-coding RNAs. Curr Opin Mol Ther 12:214–222
Marai I, El-Darawany A, Fadiel A, Abdel-Hafez M (2007) Physiological traits as affected by heat stress in sheep—a review. Small Rumin Res 71:1–12
Martinez SR, Gay MS, Zhang L (2015) Epigenetic mechanisms in heart development and disease. Drug Discov Today 20:799–811. https://doi.org/10.1016/j.drudis.2014.12.018
Mashaly M, Hendricks G 3rd, Kalama M, Gehad A, Abbas A, Patterson P (2004) Effect of heat stress on production parameters and immune responses of commercial laying hens. Poult Sci 83:889–894
Meng X, Yang J, Dong M, Zhang K, Tu E, Gao Q, Chen W, Zhang C, Zhang Y (2016) Regulatory T cells in cardiovascular diseases. Nat Rev Cardiol 13:167–179
Moraes VMB, Malheiros RD, Bruggeman V, Collin A, Tona K, Van As P, Onagbesan OM, Buyse J, Decuypere E, Macari M (2003) Effect of thermal conditioning during embryonic development on aspects of physiological responses of broilers to heat stress. J Therm Biol 28:133–140, ISSN 0306-4565. https://doi.org/10.1016/S0306-4565(02)00049-9
Nishimura D (2004) A view from the web. BioCarta. www.biocarta.com. Biotech Softw Internet Rep 2:117–120
Orozco LD, Morselli M, Rubbi L, Guo W, Go J, Shi H, Lopez D, Furlotte NA, Bennett BJ, Farber CR, Ghazalpour A, Zhang MQ, Bahous R, Rozen R, Lusis AJ, Pellegrini M (2015) Epigenome-wide association of liver methylation patterns and complex metabolic traits in mice. Cell Metab 21:905–917
Phillips C, Piggins D (1992) Farm animals and the environment. CAB International, Wallingford ISBN: 0851987885
Renaudeau D, Collin A, Yahav S, De Basilio V, Gourdine J, Collier R (2012) Adaptation to hot climate and strategies to alleviate heat stress in livestock production. Animal 6:707–728. https://doi.org/10.1017/S1751731111002448
Sahar S, Sassone-Corsi P (2013) The epigenetic language of circadian clocks. Handb Exp Pharmacol 217:29–44. https://doi.org/10.1007/978-3-642-25950-0_2
Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T (2003) Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res 13:2498–2504
Silanikove N (1992) Effects of water scarcity and hot environment on appetite and digestion in ruminants: a review. Livest Prod Sci 30:175–194
Srikanth K, Kwon A, Lee E, Chung H (2017) Characterization of genes and pathways that respond to heat stress in Holstein calves through transcriptome analysis. Cell Stress Chaperon 22:29–42
Szklarczyk D, Franceschini A, Wyder S, Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos A, Tsafou KP, Kuhn M, Bork P, Jensen LJ, von Mering C (2015) STRING v10: protein-protein interaction networks, integrated over the tree of life. Nucleic Acid Res 43:D447–D542
Szklarczyk D, Santos A, von Mering C, Jensen LJ, Bork P, Kuhn M (2016) STITCH 5: augmenting protein-chemical interaction networks with tissue and affinity data. Nucleic Acid Res 44:D380–D384
Szklarczyk D, Morris JH, Cook H, Kuhn M, Wyder S, Simonovic M, Santos A, Doncheva NT, Roth A, Bork P, Jensen LJ, von Mering C (2017) The STRING database in 2017: quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acid Res 45:D362–D368
Te Pas MFW, Wijnberg ID, Hoekman AJW, De Graaf-Roelfsema E, Keizer HA, Van Breda E, Ducro B, Van der Kolk JH (2013) Skeletal muscle transcriptome profiles related to different training intensities and detraining in Standardbred horses: a search for overtraining biomarkers. Vet J 197:717–723. https://doi.org/10.1016/j.tvjl.2013.03.052
Thornton PK, van de Steeg J, Notenbaert A, Herrero M (2009) The impacts of climate change on livestock and livestock systems in developing countries: a review of what we know and what we need to know. Agric Syst 101:113–127. https://doi.org/10.1016/j.agsy.2009.05.002
Tidball JG (2017) Regulation of muscle growth and regeneration by the immune system. Nat Rev Immunol 17:165–178. https://doi.org/10.1038/nri.2016.150
Unternaehrer E, Luers P, Mill J, Dempster E, Meyer AH, Staehli S, Lieb R, Hellhammer DH, Meinlschmidt G (2012) Dynamic changes in DNA methylation of stress-associated genes (OXTR, BDNF ) after acute psychosocial stress. Transl Psychiatry 2:e150. https://doi.org/10.1038/tp.2012.77
Veldman JE, Roord JJ, O'connor AF, Shea JJ (1984) Autoimmunity and inner ear disorders: an immune-complex mediated sensorineural hearing loss. Laryngoscope 94:501–507. https://doi.org/10.1288/00005537-198404000-00014
Wang SH, Cheng CY, Tang PC, Chen CF, Chen HH, Lee Y-P, Huang S-Y (2015) Acute heat stress induces differential gene expressions in the testes of a broiler-type strain of Taiwan country chickens. PLoS One 10(5):e0125816. https://doi.org/10.1371/journal.pone.0125816
Weinberg MS, Morris KV (2016) Transcriptional gene silencing in humans. Nucleic Acid Res 44:6505–6517. https://doi.org/10.1093/nar/gkw139
West J (2003) Effects of heat-stress on production in dairy cattle. J Dairy Sci 86:2131–2144
Yahav S, Goldfeld S, Plavnik I, Hurwitz S (1995) Physiological responses of chickens and turkeys to relative humidity during exposure to high ambient temperature. J Therm Biol 20:245–253
Yossifoff M, Kisliouk T, Meiri N (2008) Dynamic changes in DNA methylation during thermal control establishment affect CREB binding to the brain-derived neurotrophic factor promoter. Eur J Neurosci 28:2267–2277. https://doi.org/10.1111/j.1460-9568.2008.06532.x
Young RA (1990) Stress proteins and immunology. Ann Rev Immunol 8:401–420
Zahoor I, Mitchell MA, Hall S, Beard PM, Gous RM, De Koning DJ, Hocking PM (2016) Predicted optimum ambient temperatures for broiler chickens to dissipate metabolic heat do not affect performance or improve breast muscle quality. Br Poult Sci 57:134–141. https://doi.org/10.1080/00071668.2015.1124067
Zahoor I, de Koning DJ, Hocking PM (2017) Transcriptional profile of breast muscle in heat stressed layers is similar to that of broiler chickens at control temperature. Genet Sel Evol 49:69. https://doi.org/10.1186/s12711-017-0346-x
Zhang L, Yue HY, Zhang HJ, Xu L, Wu SG, Yan HJ, Gong YS, Qi GH (2009) Transport stress in broilers: I. Blood metabolism, glycolytic potential, and meat quality. Poult Sci 88:2033–2041. https://doi.org/10.3382/ps.2009-00128
Funding
This study was carried out with the support of “Cooperative Research Program for Agriculture Science & Technology Development (Project title: Biomarker investigation for livestock animal productivity through combinatory analysis with big-data related to genetic elements affecting economic traits, Project No. PJ011876; Project title: Investigation of expression profiles and genetic networks related to heat stress for chicken, Project No. PJ011221)” Rural Development Administration, Republic of Korea, and this study was supported by 2017 the RDA Fellowship Program of National Institute of Animal Science, Rural Development Administration, Republic of Korea.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
Normalized heart tissue gene expression data (in FPKM). (XLSX 4098 kb)
ESM 2
Normalized muscle tissue gene expression data (in FPKM). (XLSX 3884 kb)
ESM 3
Normalized spleen tissue gene expression data (in FPKM). (XLSX 4208 kb)
ESM 4
Functional annotation analysis of the differentially expressed genes in the experimental groups at different times of the day using the DAVID software. (XLSX 30 kb)
ESM 5
Overview of network analysis. The data were obtained from protein network analysis using the STRING software, Protein and metabolite analysis using the STITCH software, and grouped pathways analysis using the Cytoscape software with the ClueGo app. (XLSX 141 kb)
ESM 6
Network analysis figures. All figures were converted to pdf files. (ZIP 47344 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
te Pas, M.F.W., Park, W., Srikanth, K. et al. Transcriptomic profiles of muscle, heart, and spleen in reaction to circadian heat stress in Ethiopian highland and lowland male chicken. Cell Stress and Chaperones 24, 175–194 (2019). https://doi.org/10.1007/s12192-018-0954-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12192-018-0954-6