
Abstract
Soil microbial biomass is an important indicator to measure the dynamic changes of soil carbon pool. It is of great significance to understand the dynamics of soil microbial biomass in plantation for rational management and cultivation of plantation. In order to explore the temporal dynamics and influencing factors of soil microbial biomass of Keteleeria fortunei var. cyclolepis at different stand ages, the plantation of different ages (young forest, 5 years; middle-aged forest, 22 years; mature forest, 40 years) at the Guangxi Daguishan forest station of China were studied to examine the seasonal variation of their microbial biomass carbon (MBC) and microbial biomass nitrogen (MBN) by chloroform fumigation extraction method. It was found that among the forests of different age, MBC and MBN differed significantly in the 0–10 cm soil layer, and MBN differed significantly in the 10–20 cm soil layer, but there was no significant difference in MBC for the 10–20 cm soil layer or in either MBC or MBN for the 20–40 cm soil layer. With increasing maturity of the forest, MBC gradually decreased in the 0–10 cm soil layer and increased firstly and then decreased in the 10–20 cm and 20–40 cm soil layers, and MBN increased firstly and then decreased in all three soil layers. As the soil depth increased, both MBC and MBN gradually decreased for all three forests. The MBC and MBN basically had the same seasonal variation in all three soil layers of all three forests, i.e., high in the summer and low in the winter. Correlation analysis showed that MBC was significantly positively correlated with soil organic matter, total nitrogen, and soil moisture, whereas MBN was significantly positively correlated with soil total nitrogen. It showed that soil moisture content was the main factor determining the variation of soil microbial biomass by Redundancy analysis. The results showed that the soil properties changed continuously as the young forest grew into the middle-aged forest, which increased soil microbial biomass and enriched the soil nutrients. However, the soil microbial biomass declined as the middle-age forest continued to grow, and the soil nutrients were reduced in the mature forest.
Similar content being viewed by others


Introduction
The soil microbial biomass regulates the carbon and nitrogen cycle in terrestrial ecosystems and also plays a key role in the conversion and supply of nutrients (Berg and Smalla 2009). To ensure effective management and cultivation of plantation, it is imperative to thoroughly understand the dynamic changes of soil microbial biomass and its influencing factors during forest growth. The soil microbial biomass usually exhibits seasonal variation because it is strongly affected by water, temperature, soil physicochemical properties, nutrients, etc., all of which are in turn highly seasonal (Yao et al. 2011; Wang et al. 2016). Previous studies on the seasonal variation of soil microbial biomass in plantation mainly focused on the differences in land use pattern (Pandey et al. 2010; Li et al. 2018), forest type (Ravindran and Yang 2015), and forest age (Wen et al. 2014). For plantation of different age, existing studies on the soil microbial biomass have studied the surface soil layer (< 20 cm) of broad-leaved forests in tropical, subtropical, and temperate areas as well as of coniferous forests in subpolar areas (Yang et al. 2010; Vidyanagar 2010), but research is lacking on the soil microbial biomass of coniferous forests in subtropical regions or of deep soil layers.
In China, Guangxi is a major area for the production of high-valued tropical and subtropical tree species. Although the local geographical conditions are particularly suitable for the cultivation of high-valued tree species, the development of high-valued tree species is presently lacking. The three fast-growing tree species (Pinaceae, Cunninghamia, Eucalyptus) are the dominant species, but the potential of forest lands remains to be further exploited since other trees are largely neglected. For the sake of local economy in the mountainous areas of Guangxi, it is very important to make full use of the ecological advantages of Guangxi (in both tree species resources and geographical environment) and actively develop plantation of high-valued tree species to improve the productivity of forest land. Keteleeria fortunei var. cyclolepis is a high-valued native tree species in Guangxi that belongs to the Keteleeria genus of the Pinaceae family. It has very high economic value because it is an excellent species for both landscaping and the production of precious timber, and it is also an ideal species for reforestation in mountains (Liu et al. 2017). Present research on the artificial forest of K. fortunei var. cyclolepis focused mostly on the effects of topography, meteorology, and habitat on the forest growth and biomass, as well as the physiochemical properties of the forest soil (Huang et al. 2016), and there is currently no published work on the soil microbial biomass.
In this paper, seasonal dynamics of soil microbial biomass in K. fortunei var. cyclolepis plantation forests of different age at the Guangxi Daguishan forest station was studied to understand whether the changes of soil microbial biomass during the growth of K. fortunei var. cyclolepis plantation changed its soil properties. The soil fertility status was analyzed, and the relevant factors affecting the soil microbial biomass were discussed. The results provide a scientific basis for the rational management of K. fortunei var. cyclolepis plantation as well as cultivation of other coniferous forests in tropical and subtropical regions.
Materials and methods
Study site
The test sites (111° 20′ 5′′ E, 24° 14′ 25′′ N) are located at the Liupai branch of the Daguishan forest station in the northeast to the Hezhou city of Guangxi, China. The landform consists of low hills and low mountains, and the elevation is 80–1204 m. The test sites have humid subtropical monsoon climate with an average annual temperature of 19.3 °C, an average annual rainfall of 2056 mm, and an average relative humidity of 82% (Liu et al. 2017). The soil is developed from residual parent material and consists mainly of mountainous brow soil. The soil texture is clay. The soil thickness is about 90–120 cm. The selected test sites have essentially uniform environmental conditions and management measures. The age of the artificial K. fortunei var. cyclolepis forests varies from 5 years (young forest) to 22 years (middle-aged forest) and 40 years (mature forest). Additional information of the test sites is given in Table 1.
Methods
Three replicate plots (20 m × 20 m) were randomly selected in each forest site, and soil samples were taken at the nine plots from soil layers at 0–10, 10–20, and 20–40 cm in March, June, September, and December of 2016, hence each time 27 samples were brought back to the laboratory for analysis. Each fresh soil sample was divided into two parts. The first half was passed through a stainless steel filter (2 mm) and measured for soil microbial carbon (MBC) and soil microbial nitrogen (MBN), and the other half was air-dried, sifted, and measured for soil organic carbon (SOC), total nitrogen (TN), total phosphorus (TP), pH, bulk density (BD), and soil water content (SWC) (Oksanen 2001; Binkley and Fisher 2012).The MBC and MBN were measured by chloroform fumigation (Vance et al. 1987). Soil was weighed (10.00 g × 4) to give four portions, which were placed in a vacuum dryer containing chloroform and incubated at 25 °C for 24 h. The soil samples before and after fumigation were mixed with aqueous K2SO4 (0.5 mol/L, 40 mL), shaken at 300 rpm for 30 min, and sifted. The filtrate was measured on a multi N/C 3100 (Analytik Jena AG, Germany) instrument.
Soil organic carbon was determined by combustion method and multi N/C 2100 analyzer (Analytik Jena AG, Jena German). Soil total nitrogen was determined by Kjeldahl method using Automatic kjeldahl apparatus (Kjeltec 8400, Foss, Hiller d, Danmark). The content of phosphorus was determined by molybdenum-antimony colorimeter using uv-2600 spectrophotometer. The soil pH was determined by the glass electrode method. The soil bulk density was calculated by dividing the dry soil weight by the volume of the ring knife used for sampling (Binkley and Fisher 2012).
Data analysis
The seasonal variation of the soil physicochemical properties and microbial biomass of the artificial K. fortunei var. cyclolepis forests of different age was analyzed and plotted with Excel 2007. Variance analysis, Pearson correlation analysis, and Duncan’s multiple range test were conducted with SPSS19.0. The significance level for all statistical analyses was set at α = 0.05. In order to reflect the effect of soil physical and chemical properties on soil microbial biomass, the data of soil microbial biomass and soil physical and chemical properties were sequenced and made redundancy analysis (RDA) by Canoco 5.0 software.
Results
Vertical variation of soil microbial biomass and physicochemical properties
Figure 1 shows the vertical variation of the soil physicochemical properties and the microbial biomass. For the 0–10 cm soil layer, significant difference among forests of different age was found (P < 0.05) for TN, TP, MBC, and MBN, but not (P > 0.05) for SOC, pH, BD, SWC, and MBC/MBN. For the 10–20 cm soil layer, significant difference among forests of different age was found for SOC, TN, MBC, and MBC/MBN, but did not exist for MBN, TP, pH, BD, and SWC. For the 20–40 cm soil layer, there was no significant difference among forests of different age for MBC, MBN, MBC/MBN, SOC, TN, TP, pH, BD, and SWC.

The vertical changes in soil microbial biomass and physicochemical properties at different sites of artificial K. fortunei var. cyclolepis forests. Different capital letters of the same site indicate significant differences between soil layers, and different lower case letters of the same soil layer indicate significant differences across forest sites
In general, for the soil of all three forest sites, SOC, TN, TP, SWC, MBC, MBN gradually decreased with soil depth, BD increased with soil depth, and pH and MBC/MBN did not seem to clearly vary with soil depth. For the young forest, there was extremely significant difference among soil layers for SOC, TN, and TP but no significant difference for pH, BD, and SWC. For the middle-aged forest, there was extremely significant difference among soil layers for SOC, TN, and TP, significant difference for SWC, and no significant difference for pH and BD. For the mature forest, there was extremely significant difference among soil layers for SOC, TN, and TP, but no significant difference for pH, BD, and SWC.
Seasonal variation of soil microbial biomass
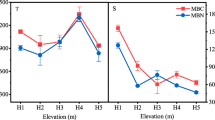
Figure 2 shows the seasonal variation of the soil microbial biomass for the artificial K. fortunei var. cyclolepis forests of different age. It can be noted that for all soil layers of all forest sites, both MBC and MBN were the highest in summer and the lowest in winter. For the 0–10 cm soil, MBC and MBN fell in the order of summer > fall > spring > winter. For the 20–40 cm soil, MBC and MBN fell in the order of summer > spring > fall > winter. For the 10–20 cm soil, except the MBC of the young forest showed an order of summer > spring > fall > winter, the MBC of the middle-aged and mature forests, as well as the MBN of all three forest, all showed the order of summer > fall > spring > winter.

The seasonal variation in soil microbial biomass at different forest sites of K. fortunei var. cyclolepis (Mean ± SE)
The seasonal variation of MBC/MBN was different. For the 0–10 cm soil, the MBC/MBN of all three forests fell in the order of spring > summer > winter > fall. For the 10–20 cm soil, the MBC/MBN of the young forest fell in the order of spring > winter > fall > summer, and the MBC/MBN of the middle-aged and mature forests fell in the order of spring > fall > winter > summer. For the 20–40 cm soil, the MBC/MBN of young and middle-aged forests fell in the order of spring > summer > winter > fall, and the MBC/MBN of the mature forest fell in the order of winter > spring > fall > summer.
Influence factors for the soil microbial biomass
Pearson correlation analysis (Table 2) showed that for the 0–10 cm soil, significant positive correlation existed between MBC and SOC, TN, SWC as well as between MBC/MBN and SOC, and there was also extremely significant positive correlation between MBN and TN. For the 10–20 cm soil, there were extremely significant positive correlation between MBC and SOC, significant positive correlation between MBN and TN, significant positive correlation between MBC/MBN and SOC, and significant negative correlation between MBC/MBN and TN. For the 20–40 cm soil, significant positive correlation existed between MBN and TN.
According to the RDA ordination diagram (Fig. 3), the total explanatory rate of soil physicochemical properties to microbial biomass change is 97.3%, the explanatory rates of axis I and axis II are 91.42% and 5.69%, respectively, and the cumulative explanatory rate is 97.11%, accounting for 98.8% of the total explanatory rate. It showed that the first two axes could reflect the relationship between soil microbial biomass and physicochemical properties well. After gradual selection of SWC, SOC, TN, pH, BD and TP, the cumulative values of explanatory variables were 75.6%, 94.2%, 95.3%, 96.5%, 97.2% and 97.3%, indicating that SWC was the main factor determining the variation of soil microbial biomass, and its explanatory rate was 75.6%.

RDA ordination diagram of soil microbial biomass and physicochemical properties
Discussions
Difference in the soil microbial biomass of different forests
The results showed that there was significant differences in soil microbial biomass between different forest ages in the 0–20 cm soil layer. With the increase of soil depth, there were no significant differences in soil microbial biomass between different forest ages, and both MBC and MBN gradually decreased. It indicated that the microbial biomass was more enriched and had greater diversity in the surface soil layers (0–10 and 10–20 cm) than in the deep soil layer (20–40 cm) for the K. fortunei var. cyclolepis plantations of different ages. It can be seen that the forest age strongly affected the soil microbial biomass in the surface layers. It is consistent with the research results of Xu et al. (2013) and Wen et al. (2014). However, the study of An et al. (2011) showed that the number of microorganisms was the highest in the soil layer of 10–40 cm, which maybe because the soil in the study area was sandy loam, and water and nutrients were easily diffused and leached to the deep layer, leading to the enrichment of microorganisms in the deep layer. The study area belongs to dark brown forest soil with more inputs of fine roots and litters and better ventilation and permeability in the surface soil, which was conducive to the growth and reproduction of microorganisms. On the other hand, as the soil layer deepened, soil water content decreased and the soil porosity declined which reduced the ion exchange in soil, impeded the diffusion of nutrients, and impaired the growth and distribution of soil microbes. There were no significant differences in the nutrients and physicochemical properties of soil among forests of different age, and thus their soil microbial biomass could not be well distinguished from each other (Fierer et al. 2003; Stockdale and Brookes 2006).
Generally speaking, as the age of the forest increased, the soil microbial biomass increased firstly and then decreased, in the order of middle-aged forest > young forest > mature forest. This was in line with the results reported by Wu et al. (2016) on the soil microbes of the Larix principis-rupprechtii Mayr forests at the Shanxi Taiyue mountain. It could be speculated that the young forest of K. fortunei var. cyclolepis had smaller trees, and forest floor light, humidity, and temperature were all favorable to the metabolism and growth of soil microbes. As the forest age increased, the amount of litter increased, which enriched the soil nutrients and provided additional energy source for microbial metabolism and synthesis. However, when the forest matured further, the canopy of the forests became denser, which reduced forest floor light and slowed decomposition of litter. Vegetations such as shrubs and herbs also declined consequently, which decreased soil nutrients and together lead to slower microbial growth (Kaiser et al. 2011; Song et al. 2015).
The ratio of soil microbial carbon to soil microbial nitrogen is often used as an important indicator of changes in the microbial community, and higher ratio indicates higher fungal biomass (Zhou and Wang 2015). In this study, there was no significant difference in MBC/MBN in soil layers of 0–10 cm and 20–40 cm between different forest ages, while MBC/MBN in soil layers of 10–20 cm was significantly different between different forest ages, and MBC/MBN in soil of young and medium forests was significantly greater than that in forest (Fig. 2). It indicated that K. fortunei var. cyclolepis plantations had little effect on the microbial community in the soil surface (0–10 cm) and deep (20–40 cm), but greater effect on the microbial community in the soil middle (10–20 cm). It may be that the soil organic matter was beneficial to the growth of soil fungal growth, while total nitrogen content was beneficial to the growth of the soil bacteria. There was significant differences between soil organic matter and total nitrogen content in 10–20 cm soil layer of different forest ages, which causes the difference of MBC/MBN in the soil layer of different forest ages. The specific i mechanism and reason of the impact still needed further in-depth study (Trap et al. 2011). With the deepening of soil layer, the MBC/MBN of K. fortunei var. cyclolepis plantation increased gradually, which similar to the previous research results (Fierer et al. 2003; An et al. 2011). It mainly because the soil surface nutrient limit was lower and it was more suitable for bacteria to survive.
Seasonal variation and influence factors of soil microbial biomass
The seasonal soil microbial biomass is strongly affected by humidity, temperature, soil physicochemical properties, and other environmental factors (Classen et al. 2007; Freppaz et al. 2014). In the current study of the artificial K. fortunei var. cyclolepis forests, the observed soil microbial carbon (MBC) and soil microbial nitrogen (MBN) were clearly high in the summer and low in the winter. That is, MBC and MBN were high in the growth season and low in the hibernation season. Similar trend was also observed by Devi and Yadava (2006) for the mixed forest of Quercus Linn in Manipur, India, by Ruan et al. (2004) for tropical rainforest, as well as by Li et al. (2014) for the mixed forest of Cunninghamia lanceolata Hook, Pinus massoniana Lamb and Lithocarpus glaber Nakai in the hilly areas of central Hunan in China. The higher temperature in the summer promotes the decomposition of litter, and nutrients and carbohydrates are brought into the soil from the earth surface by rain water, which accelerates the growth of soil microbes and increases both MBC and MBN. Conversely, since the winter is colder and drier, the decomposition of litter is slower and both microbial growth and activity are inhibited, leading to lower MBC and MBN (Lipson and Schmidt 2000). The seasonal variation of soil MBC/MBN in K. fortunei var. cyclolepis plantations of different forest ages also showed a certain rule, which was mainly manifested as that the soil MBC/MBN of the three forest ages was the largest in spring, and the soil MBC/MBN of the young and middle forests was greater than that of the mature forests, especially in the soil layer of 10–40 cm. We speculated that this may be caused by the increase of external temperature in spring, the smaller canopy density in young and middle forests, and the more sensitive response of surface soil to external temperature and the more light it received, which is conducive to fungus breeding and growth (Kaiser et al. 2014; Jefferies et al. 2010).
Soil microbial biomass is related to environmental factors such as temperature, humidity, soil moisture, soil nutrients, soil physicochemical properties, etc. (Devi and Yadava 2006; Liu et al. 2014). Current results showed that in the plantation of K. fortunei var. cyclolepis, MBC was significantly positively correlated with soil organic matter, total nitrogen, and soil moisture, while MBN was significantly positively correlated with soil total nitrogen. Therefore, soil organic matter and total nitrogen were important influence factors of MBC and MBN. High soil organic matter and total nitrogen can provide the soil microbes with sufficient carbon, nitrogen, and energy source for their synthesis and metabolism (Li et al. 2014). These results are consistent with previous findings. Yang et al. (2009) studied the seasonal changes of soil microbial biomass in the plantation of Larix gmelinii Kuzen and found significantly positive correlation between soil microbial biomass (both carbon and nitrogen) and soil organic matter and total nitrogen. The global analysis by Xu et al. (2013) on the soil microbial biomass (carbon, nitrogen, and phosphorus) in terrestrial ecosystems demonstrated that the spatial pattern of soil microbial biomass (carbon and nitrogen) was consistent with that of soil organic matter and total nitrogen.
In conclusion, the soil nutrients in middle age were more nutritious than young and mature forests of K. fortunei var. cyclolepis plantations, that is to say, the soil nutrients gradually increased with the development of the plantations, and reached the maximum at middle age. And then the soil nutrients gradually decreased with the increase of forest age, while the soil fertility declines also.
References
An R, Gong JR, You X, Ge ZW, Duan QW, Yan X (2011) Seasonal dynamics of soil microorganisms and soil nutrients in fast-growing populus plantation forests of different ages 4in Yili, Xinjiang, China. Chin J Plant Ecol 35(4):389–401 (in Chinese)
Berg G, Smalla K (2009) Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol Ecol 68(1):1–13
Binkley D, Fisher R (2012) Ecology and management of forest soils. Wiley, New York, pp 151–153
Classen AT, Overby ST, Hart SC, Koch GW, Whitham TG (2007) Season mediates herbivore effects on litter and soil microbial abundance and activity in a semi-arid woodland. Plant Soil 295(2):217–227
Devi NB, Yadava PS (2006) Seasonal dynamics in soil microbial biomass C, N and P in a mixed-oak forest ecosystem of Manipur, North-east India. Appl Soil Ecol 31(3):220–227
Fierer N, Schimel JP, Holden PA (2003) Influence of drying-rewetting frequency on soil bacterial community structure. Microb Ecol 45(1):63–71
Freppaz M, Said-Pullicino D, Filippa G, Curtaz F, Celi L, Zanini E (2014) Winter-spring transition induces changes in nutrients and microbial biomass in mid-alpine forest soils. Soil Biol Biochem 78:54–57
Huang RL, He YH, Jiang Y, Zhang JC, Liu F, Jiang Y (2016) Relationships between meteorological factors and artificial forest growth of Keteleeria cyclolepis Flous in Guangxi. Guangxi For Sci 45(3):328–333
Jefferies RL, Walker NA, Edwards KA, Dainty J (2010) Is the decline of soil microbial biomass in late winter coupled to changes in the physical state of cold soils? Soil Biol Biochem 42(2):129–135
Kaiser C, Fuchslueger L, Koranda M, Gorfer M, Stange CF, Kitzler B, Rasche F, Strauss J, Sessitsch A, Boltenstern SZ, Richter A (2011) Plants control the seasonal dynamics of microbial N cycling in a beech forest soil by belowground allocation. Ecology 92(5):1036–1051
Kaiser C, Franklin O, Dieckmann U, Richter A (2014) Microbial community dynamics alleviate stoichiometric constraints during litter decay. Ecol Lett 17(6):680–690
Li SL, Fang X, Xiang WH, Sun WJ, Zhang SJ (2014) Soil microbial biomass carbon and nitrogen concentrations in four subtropical forests in hilly region of central Hunan province, China. Sci Silva Sin 50(5):8–16
Li Y, Zhang LP, Fang SZ, Tian Y, Guo J (2018) Variation of soil enzyme activity and microbial biomass in poplar plantations of different genotypes and stem spacings. J For Res 29(4):963–972
Lipson DA, Schmidt SK, Monson ARK (2000) Carbon availability and temperature control the post-snowmelt decline in alpine soil microbial biomass. Soil Biol Biochem 32(4):441–448
Liu D, Fang SZ, Tian Y, Dun XJ (2014) Seasonal and clonal variations of microbial biomass and processes in the rhizosphere of poplar plantations. Appl Soil Ecol 78:65–72
Liu XS, Jiang Y, Huang RL, Liu F, Xiao YF, Jiang Y, Wei SX (2017) Variation in traits of cone and seed of Keteleeria fortunei var. cyclolepis and its relationship with environmental factors. Guihaia 37(1):118–126 (in Chinese)
Oksanen L (2001) Logic of experiments in ecology: is pseudoreplication a pseudoissue? Oikos 94(1):27–38
Pandey CB, Singh GB, Singh SK, Singh RK (2010) Soil nitrogen and microbial biomass carbon dynamics in native forests and derived agricultural land uses in a humid tropical climate of India. Plant Soil 333(1):453–467
Ravindran A, Yang SS (2015) Effects of vegetation type on microbial biomass carbon and nitrogen in subalpine mountain forest soils. J Microbiol Immunol 48(4):362–369
Ruan HH, Zou XM, Scatena FN, Zimmerman JK (2004) Asynchronous fluctuation of soil microbial biomass and plant litterfall in a tropical wet forest. Plant Soil 260(1):147–154
Song P, Ren H, Jia Q, Guo J, Zhang N, Ma K (2015) Effects of historical logging on soil microbial communities in a subtropical forest in southern China. Plant Soil 397(2):115–126
Stockdale EA, Brookes PC (2006) Detection and quantification of the soil microbial biomass impacts on the management of agricultural soils. J Agric Sci 144(4):285–302
Trap J, Laval K, Akpa-Vinceslas M, Gangneux C, Bureau F, Decaëns T, Aubert M (2011) Humus macro-morphology and soil microbial community changes along a 130-yr-old Fagus sylvatica, chronosequence. Soil Biol Biochem 43(7):1553–1562
Vance ED, Brookes PC, Jenkinson DS (1987) An extraction method for measuring soil microbial biomass C. Soil Biol Biochem 19(6):703–707
Vidyanagar V (2010) Seasonal and temporal variation in soil microbial biomass C, N and P in different types land uses of dry deciduous forest ecosystem of Udaipur, Rajasthan. Appl Ecol Environ Res 8(4):377–390
Wang X, Xu L, Wan R, Chen Y (2016) Seasonal variations of soil microbial biomass within two typical wetland areas along the vegetation gradient of Poyang Lake, China. Catena 137:483–493
Wen L, Lei P, Xiang W, Yan W, Liu S (2014) Soil microbial biomass carbon and nitrogen in pure and mixed stands of Pinus massoniana, and Cinnamomum camphora, differing in stand age. Forest Ecol Manag 328:150–158
Wu R, Kang FF, Han HR, Cheng XQ, Zhou WS, Wang LX, Chen J, Tian P (2016) Soil microbial properties in Larix principis-rupprechtii plantations of different ages in Mt. Taiyue, Shanxi, China. Chin J Ecol 35(12):3183–3190
Xu X, Thornton PE, Post WM (2013) A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Glob Ecol Biogeogr 22(6):737–749
Yang K, Zhu JJ, Zhang JX, Yan QL (2009) Seasonal dynamics of soil microbial biomass C and N in two larch plantation forests with different ages in Northeastern China. Acta Ecol Sin 29(10):5500–5507 (in Chinese)
Yang K, Zhu J, Zhang M, Yan Q, Sun J (2010) Soil microbial biomass carbon and nitrogen in forest ecosystems of Northeast China: a comparison between natural secondary forest and larch plantation. J Plant Ecol 3(3):175–182
Yao H, Bowman D, Shi W (2011) Seasonal variations of soil microbial biomass and activity in warm- and cool-season turfgrass systems. Soil Biol Biochem 43(7):1536–1543
Zhou ZH, Wang CK (2015) Soil resources and climate jointly drive variations in microbial biomass carbon and nitrogen in China’s forest ecosystems. Biogeosciences 12(14):11191–11216
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: The project was fully funded by Guangxi Key Research and Development Plan (NO. AB163800100) and Forestry science and technology programs of Guangxi (NO.32 GuiLlinkezi [2016]).
The online version is available at http://www.springerlink.com
Corresponding editor: Zhu Hong.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Wang, Y., Liu, X., Chen, F. et al. Seasonal dynamics of soil microbial biomass C and N of Keteleeria fortunei var. cyclolepis forests with different ages. J. For. Res. 31, 2377–2384 (2020). https://doi.org/10.1007/s11676-019-01058-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-019-01058-w