
Abstract
This study was aimed at evaluating the effect of a microbial pest-controlling product (MPCP) with the active substance Bacillus thuringiensis ssp. aizawai (strain: ABTS-1857) on adults and larvae of honeybees. To determine the contamination levels of Bt spores in different matrices, a colony-feeding study under semi-field conditions was performed. Furthermore, two chronic adult trials and a chronic larval study were conducted under laboratory conditions to test the effects of different concentrations of the plant protection product (PPP) on the development and mortality. Possible modifications of the chronic oral toxicity test were assessed by additional pollen feeding. Our results showed that Bt spores were detected in all matrices over the entire test duration in different concentrations, decreasing over time. The survival of adult bees and larvae was negatively affected in laboratory conditions after a chronic exposure to the MPCP depending on the tested concentrations. Moreover, the earliest sign of bee mortality, resulting from exposure to ABTS-1857, was recorded only after 96 h at the highest tested concentration. Pollen feeding to adults significantly increased the survival of the treated bees. In conclusion, the PPP with the Bt strain ABTS-1857 showed an effect on the mortality of adults and larvae under laboratory conditions. Further studies with Bt-based PPPs under realistic field conditions are necessary to evaluate the potential risk of those MPCPs on honeybees.
Similar content being viewed by others


Introduction
Recently, increasing numbers of studies have been published regarding the potential adverse effects of chemical plant protection products (PPPs) on insect pollinators including Apis and non-Apis bees. To overcome the harmful side effects of those PPPs on non-target organisms, biological methods for pest control have been developed. One approach is the use of microbial pest-controlling products (MPCPs), which seem to be highly specific for particular insect orders (Schünemann et al. 2014). During the last decades, a wide range of microbial pest controlling products (MPCPs) has been developed (Köhl et al. 2019). Isolates of entomopathogenic bacteria, such as Bacillus thuringiensis (Bt), have been used commercially to control agricultural and forest insect pests (Bravo et al. 2011). Because of the selective impacts on insects of the orders Lepidoptera, Coleoptera, and Diptera as well as the presumed safety of non-target insects (Schnepf et al. 1998), Bt is applied in integrated pest management (IPM) in organic farming and is often combined with beneficial insects, such as pollinators (Dietrich et al. 2014). The toxic action of Bt-based products in the susceptible insects occurs after the uptake in the gut during the sporulation phase of the bacteria, when particularly insecticidal toxins, such as Cry and Cyt toxins, are produced as crystal inclusions (Bravo et al. 2007; Pardo-López et al. 2013).
Despite the presumed selective action, some Cry toxins have been found to be toxic for other insect groups as well (Feitelson et al. 1992; Feitelson 1993; Schnepf et al. 1998). In particular, a Hymenopteran-specific effect has to be excluded because of the common use with different bioagents (Dietrich et al. 2014). During their foraging activity, bees may be exposed to the Bt products either acutely after spray application or chronically by ingestion of contaminated pollen and nectar; thus, the products can be assumed to be transported back to the colony, leading to exposure of in-hive bees and stored matrices. Furthermore, a long-term effect due to the persistence of Bt sprays is possible. Although a fast reduction of Bt population and toxicity in the environment after application is reported (Schnepf et al. 1998; Haddad et al. 2005; Dietrich et al. 2014), the Bt spores still survive several years after use (Addison 1993). However, there is no information about the viability of Bt spores in the collected matrices, i.e., nectar and pollen, in the bee colony after application in different bee-attractive crops.
The results of the few studies investigating the effects of Bt on bees are somewhat inconsistent and vary with the tested strain and concentrations. D’Urso et al. (2017) reported some behavioral symptoms and physiological midgut changes after acute single exposure to Bacillus thuringiensis spp. aizawai and kurstaki (GC-91 strain). Moreover, reduced survival of honeybee workers is observed for further Bt strains (Brighenti et al. 2007; Libardoni et al. 2018; Potrich et al. 2018). However, the toxicity of MPCPs appears to depend on the tested exposure route (Soni and Thakur 2011). In contrast to exposure by feeding, some application methods, such as stripping or spraying of various MPCPs, do not affect the survival of bees under laboratory conditions (Brighenti et al. 2007; Mommaerts et al. 2010; Libardoni et al. 2018; Potrich et al. 2018). Mommaerts et al. (2010) reported 100% worker mortality of tested bumble bees after 1 week of oral exposure at the maximum field recommended rate of Bacillus thuringiensis ssp. aizawai via sugar water. On the other hand, they did not report any mortality after exposure to Bacillus thuringiensis ssp. kurstaki.
Several recent studies have focused on the interactions between single species in the microbiome of the bee gut and their community dynamics in relation to the bees’ health (Engel et al. 2016). Motta et al. (2018) showed increased mortality for bees lacking gut microbiota relative to that observed for bees with a conventional gut microbiota when they exposed to the opportunistic bacteria Serratia marcescens kz19, which is commonly detected at very low frequencies in the bee gut (Moran et al. 2012). This indicates the role of the gut microbiome as a part of the immune system of bees.
Besides using Bt as plant protection products, some products containing Bacillus thuringiensis ssp. aizawai (strain: B401) are currently used to control the greater wax moth (Galleria mellonella L.) in several countries. However, they are recommended to be applied on the storage wax combs, which can minimize the exposure of bees to the Bt spores. Vandenberg and Shimanuki (1990) reported that honey produced by bees on treated combs placed in the hive contained very low levels of viable B. thuringiensis spores after 20 weeks. Nevertheless, they did not investigate the effect on bees after inserting the treated frames.
Our aims were (1) to investigate the distribution of the Bt spores in different matrices within the honeybee colony and (2) to assess the effect of a MPCP with the active ingredient Bacillus thuringiensis ssp. aizawai (strain: ABTS-1857) on adults and larvae of A. mellifera. To our knowledge, neither the persistence and degradation of Bt in the bee colony nor a chronic larval test under laboratory conditions has ever been investigated before, but the results could be of great importance for bee health. In addition, standardized test systems for the impact of MPCPs on adults and larvae of bees have to be developed to evaluate potential risks for pollinators.
Materials and methods
Test organisms
A. mellifera
Honeybee workers and larvae were obtained from healthy colonies at the apiary of the JKI in Braunschweig, Germany. Each colony comprised about 15,000 workers and a fertile 1-year-old queen. One colony with approx. 5000 bees was placed in a tent to investigate the distribution of Bt within the bee colony under semi-field conditions.
B. thuringiensis ssp. aizawai (strain: ABTS-1857)
A registered PPP with the active substance B. thuringiensis ssp. aizawai (strain: ABTS-1857) was used. Based on the recommended field application rate, the different concentrations were chosen stepwise to cover environmentally relevant concentrations.
Experimental procedures
Determination of Bt distribution within the bee colony
To investigate the distribution of Bt within the bee colony, a preliminary semi-field feeding experiment with the Bt strain ABTS-1857 was performed in a bee colony. The maximum field recommended rate of the PPP (0.165 %) in orchard was mixed in 2 L of 50% (w/v) sucrose solution and fed to the colony using a feeding bag. The feeding solution was marked by mixing with ca. 2 g of blue food coloring (Dr. Oetker ®) to enable a targeted sampling from the combs (Fig. 1). Samples of nectar, pollen, bee larvae, and adults were taken at regular intervals (days 2, 5, 12, 19) after application.

Marked feeding solution using blue food coloring stored in the cells of the treated colony
A sample of approximately 1.0 g was homogenized in an equal amount (w/v) of sterile ddH2O. This suspension was used for serial dilution (10−1–10−6). To detect B. thuringiensis, 100 μL of each dilution (n = 3) was plated on LB agar (10.0 g casein (Roth), 5.0 g yeast extract (Bacto), 5.0 g NaCl (Roth), 15.0 g agar (Gerbu)) and incubated over night at 30 °C.
Phenotypic B. thuringiensis colonies were counted. Colonies were randomly selected for molecular biological identification as B. thuringiensis by amplification of the CryIAa region, as described previously (Ogunjimi et al. 2000). Accordingly, a single colony was resuspended in 100 μL sterile ddH2O, followed by bacterial lysis at 95 °C for 5 min. After centrifugation at 10,000×g for 5 min, 1.0 μL supernatant was used for PCR analysis with OneTaq® DNA Polymerase (NEB) on a 3Prime thermal cycler (Techne).
Adult bioassays, chronic exposure
Chronic exposure bioassays were carried out according the OECD Guideline no. 245 (OECD 2017). Brood combs with late-stage capped cells were transferred to the laboratory and placed in an incubator under dark conditions (35 ± 2 °C, RH 60 ± 5%) for 48 h until emergence.
Newly emerged workers were collected and caged in groups of ten individuals and maintained in darkness under controlled conditions (33 ± 2 °C, RH 60 ± 5%). Five concentrations of the bioagent varying from 14 to 2730 ppm were daily prepared fresh and mixed in 50% (w/v) sucrose solution. The bees were fed ad libitum with 1-mL syringes, fitted with plugs on the upper side of the cages. The consumed amount per cage, mortality, and behavioral abnormalities were recorded daily and compared to a control (only sucrose solution) and a toxic standard (dimethoate). In the second experiment, half of the bees were exposed to pollen and wax material of the initially used frame in addition to one of three concentrations of the bioagent varying from 14 to 690 ppm. For this, 8 cells containing bee bread were cut out with a scalpel and placed in one corner of the cage, so that the bees could easily reach both matrices.
Larval bioassays, repeated exposure
Larval rearing bioassays were conducted according to OECD Guidance no. 239 (OECD 2016) during June and July. In each experiment, the queens of three healthy colonies (replicates) were caged on a wax comb for approx. 24 h to lay eggs. Thereafter, first instar larvae, 24–36 h old, were grafted into 48-welled polystyrene microplates (Greiner Bio-One) lined with Brown Cell Cups. Each cell had prior been prepared with 20 μL of the artificial diet consisting of water, d-glucose, d-fructose, yeast extract, and royal jelly. Subsequently, the microplates were placed in an incubator under dark conditions at 95% R.H. and 35 °C. Two days after grafting (3-day-old larvae), the larvae were fed with 20 μL of a diet exposing them to a dose of the bioagent varying from 0.16 to 32 μg per larvae. The tested concentrations were chosen depending on the maximum detected concentration in nectar in the colony (see Fig. 1). Each group consisted of 16 larvae from each colony (16 × 3; n = 48). Larvae were fed over the next 3 days (days 4–6) with 30, 40, and 50 μL diet per individual. The humidity inside the incubator was reduced to ca. 80% on the 7th day after grafting (day 8) and to 50% on the 14th day after grafting (day 15) until emergence. Mortality and morphological differences were recorded when feeding them daily from the 3rd to the 8th day, on the 15th and on the 21st day and compared to a control (only food solution) and a toxic standard (dimethoate). Dead individuals were recorded and removed.
Statistical analysis
The survival data were analyzed using the Kaplan-Meier test to compare the differences of the survival rates between the control and treated groups over the test duration. The level of significance in all tests was set to ≤ 0.05. All statistical analyses were performed using R (R Core Team (2019)). The packages were “survminer” (Kassambara et al. 2019), “survival,” and “dplyer.” (R: a language and environment for statistical computing; R Foundation for Statistical Computing, Vienna, Austria; https://www.R-project.org/, Version 3.6.1).
Results
Distribution of Bt spores within the bee colony
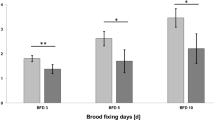
The concentration of the microbial agent B. thuringiensis ssp. aizawai, (strain: ABTS-1857) was quantified in various matrices in the bee colony over a brood cycle period by sampling the matrices nectar, bees, larvae, and pollen. Bt spores were detected in all matrices and were found over the entire sampling period in the tested colony (Fig. 2). The matrices nectar and bees contained the highest levels of Bt spores, whereas pollen and larvae showed lower levels of Bt spores. On the first sampling day, the level of Bt spores was even 85 times higher in nectar than in larvae and pollen. A reduction of the concentration was observed in nectar and bees over the experimental period of 20 days, whereas increasing Bt spore levels were found in larvae.

Number of Bt spores in CFU g−1 in the sampled matrices nectar, bees, larvae, and pollen over an experimental period of 3 weeks under semi-field conditions
Effect of chronic exposure on adult longevity
In the first experiment, a chronic exposure bioassay was conducted for 10 days to determine the effect of the tested Bt strain on the longevity of adult workers. The survival of adult bees was affected by chronic exposure to the tested product depending on the concentrations (Fig. 3; Kaplan-Meier test, p < 0.05). The earliest sign of bee mortality, resulting from exposure to ABTS-1857, was recorded only after 96 h at the highest tested concentration.

Survival rate of adults, exposed to different concentrations of the Bt strain ABTS-1857, compared to the control (C) and the toxic standard (TOX) over the test duration of 10 days (N = 6 cages/treatment, n = 10 bees/cage; Kaplan-Meier test; asterisk indicates p < 0.05)
In the second experiment, a chronic exposure bioassay was carried out with additional feeding of pollen obtained from the original colony. This additional pollen reduced the sensitivity of bees to Bt compared to bees fed only with sucrose solution (Fig. 4). Bees treated with low concentrations of ABTS-1857 (14 ppm or 140 ppm) survived the experimental period when they were fed pollen, whereas a delaying effect on the mortality was found at the higher concentration (median survival time at 690 ppm, without pollen = 5 days, with pollen = 7 days; at 10 ppm, without pollen = 8 days, with pollen > 10 days; Kaplan-Meier test, p < 0.05).

Survival rate of adults, exposed to different concentrations of the Bt strain ABTS-1857, compared to the control and the toxic standard over the test duration of 10 days with pollen (labeled as P) and without pollen (N = 3 cages/treatment, n = 10 bees/cage; Kaplan-Meier test; asterisk indicates p < 0.05)
Effect of chronic exposure on larval longevity
To evaluate the impact of the Bt strain ABTS-1857 on the longevity of larvae, a chronic Bt exposure assay was carried out during the larval stage. The development of larvae was recorded until adult emergence (21 days). The survival of larvae exposed to Bt was significantly reduced at all tested concentrations compared to the survival of untreated control larvae (Kaplan-Meier test, p < 0.005; Fig. 5). The Bt strain ABTS-1857 had a clear effect on the survival of honeybee larvae, and the major mortality occurred during the larval stages rather than in pupal stages. In all treatments except the highest concentration of the tested product and the toxic standard, pupae were able to develop until adult emergence. The larvae exposed to the toxic reference (dimethoate) showed 100% and the control 0% mortality, validating the test, according to OECD protocol no. 239.

Survival rate of larvae, exposed to different concentrations of the Bt strain ABTS-1857, compared to the control and the toxic standard over the test duration of 21 days (N = 1 well plate/treatment, n = 48 larvae/well plate; Kaplan-Meier test; asterisk indicates p < 0.05)
Discussion
Bt products are generally characterized by a low environmental persistence with a half-life period of viable Bt spores on foliage assumed to be between a few hours and 2 days (Pinnock et al. 1971; Ignoffo and Garcia 1978; Pedersen et al. 1995; Haddad et al. 2005). In contrast, we found Bt spores in all tested matrices of a bee colony at different contamination levels over the entire test period of 3 weeks. A decreasing Bt spore level over time was observed in nectar and bees. Under field conditions, the rapid degradation of Bt spores can be traced to its low UV resistance. Even under controlled greenhouse conditions in terms of temperature, humidity, and UV radiation, a high degree of degradation of Bt spores of the strain ABTS-1857 on leaves and fruits such as tomatoes was measured (Dietrich et al. 2014). Little is known about the behavior and/or vitality of the ubiquitous bacterium under bee colony conditions. Because the conditions differ substantially in the environment and in a bee colony, the persistence and activity of Bt spores depend likely on a different set of parameters, such as the absence of radiation and higher temperature and humidity. Especially temperature and humidity of bee colonies may be limiting factors for the persistence of Bt spores under colony conditions. The toxicity of Bt in orally exposed larvae of Archips xylosteana increased with rising temperatures (Li et al. 1995). Thus, temperatures of over 30 °C common in a bee colony may be beneficial for the persistence and activity of Bt. Nevertheless, MPCPs are presumed to be unstable in the colonies’ sector of the worker bee brood (Southwick and Heldmaier 1987) but quite persistent in the sector of the drone brood (Davidson et al. 2003). In contrast to the declining concentration in other matrices, the concentration of Bt in larvae increased over time in our study. This may result from a delayed use of the stored contaminated sugar solution for feeding the larvae when other food resources were absent in the tent. In the first larval stages, the brood is fed by glandular secretions of worker bees. Feeding nectar and pollen only begins at later larval stages (Winston 1987), so the oral intake of Bt via contaminated environmental matrices is expected to occur only for older larvae. Furthermore, the bacteria accumulate due to the periodic feeding of larvae and may thus cause the high mortality when no larvae survived the last sampling day in the tested colony.
The sensitivity of adult worker bees to different MPCP, especially products with Bt, was the focus of various studies (reviewed in Duan et al. 2008). While the majority of the studies showed no effect of Bt, some publications indicated a sensitivity of several bee species to particular Bt products (e.g., Libardoni et al. 2018; Mommaerts et al. 2010; Soni and Thakur 2011). Some strains, such as B. thuringiensis ssp. kurstaki (strain ABTS-351) used in the PPP Dipel®, are proven to be toxic to bees (Malone et al. 1999; Bailey et al. 2005; Brighenti et al. 2007; Mommaerts et al. 2010; Renzi et al. 2016; D’Urso et al. 2017). This indicates that the toxicity for bees depends on the Bt strain (Vandenberg and Shimanuki 1986). Furthermore, Soni and Thakur (2011) showed that the impact of MPCPs on honeybees varies with the exposure route. Bees which were orally exposed to Bt in sucrose solution, had a higher mortality rate than bees exposed by other methods, e.g., spray applications. Libardoni et al. (2018) supported this observation when comparing the effect of exposure routes with three different strains of Bt. In our study, we evaluated a possible effect of the Bt strain ABTS-1857 on adult worker bees with two chronic adult tests, based on OECD Guideline no. 245. The mortality of bees exposed to different concentrations of the PPP with the Bt strain ABTS-1857 was significantly higher than in the control bees, and the effect increased with the concentration. Additionally, the influence of colony material and/or pollen feeding seemed to decrease the sensitivity of adult worker bees when we offered them pollen cells of the original colony in our second chronic adult test. Therefore, one may assume that factors, such as contact with the bee colony and/or food quality and quantity, affect the reactions of bees on Bt. It is known that malnutrition or low food diversity not only lowers immune functions, but may also lead to increased pathogen susceptibility in bees (Alaux et al. 2010; DeGrandi-Hoffman et al. 2010). Accordingly, adequate levels of protein and a variety of pollen increase the immune competence (Alaux et al. 2010).
Under the artificial laboratory conditions in our study, larvae were highly sensitive to different concentrations of the PPP, reaching mortality rates similar to those of the toxic standard. However, larvae which had reached the pupal stage were able to develop normally. Before pupation, larvae defecate, so intestinal contents, such as food remnants or the gut microbiome, are purged (Winston 1987; Kwong and Moran 2016). As a result, accumulated populations of Bt could be removed and rendered incapable of compromising the bees’ development. Also, during the pupation time, larvae are isolated from the colony and do not contact contaminated colony material or food; thus, they are protected from exposure to entomopathogenic organisms. There is some evidence for the toxicity of the strain ABTS-1857 on other pollinator insects; e.g., feeding a PPP to Bombus terrestris induced high worker mortality and reduced the reproduction (Mommaerts et al. 2010). Because the loss of brood or young bees can have devastating consequences for a bee colony (Davis 1989), it seems worthwhile to include standardized toxicity tests of Bt strains on honeybee larvae, similar to the one for chemical PPPs reported here, in the registration process.
In view of our results and different studies, the effects of MPCPs on honeybees cannot be extrapolated from one strain to the entire group of a biologically active substance, such as Bt. Moreover, various factors influence the effect of those PPPs, such as UV radiation, humidity, and temperature. It seems essential to evaluate, modify, and adapt the current test systems used for chemical PPPs to reliably measure the potential risk of MPCPs. Adjusting the test duration is one of the necessary modifications. The earliest sign of mortality of the exposed bees under laboratory conditions was recorded only after 96 h at the highest tested concentration, additionally influenced by adult pollen feeding. An acute test with a normal test duration of 48 h or 96 h would not be able to detect these side effects and thus should be considered not sufficient for the registration process. Standardized test guidelines, as established for chemical PPPs, should be developed and applied in the risk assessment of MPCPs. Moreover, exposure routes and different environmental conditions such as the nutrition by pollen feeding appeared to affect the results, too, and have to be taken into account. The mode of action of biological PPPs under the common colony conditions, which vary significantly with the environmental conditions, needs to be investigated. Our study showed the effect of the strain ABTS-1857 on larvae and adult honeybees, but other variables, such as food quality and quantity, strain pathogenicity, weather, or race characteristics, may only be considered by examining natural interactions under field conditions (Duan et al. 2008; Soni and Thakur 2011). The most recent studies on the role of the gut microbiome of honeybees on bee health guide toward further studies determining the impact of MPCPs on the composition and development of the gut microbiome of honeybees. Thorough examinations of the effect of Bt and other MPCPs on the honeybee and other bee species under field conditions will help to understand the natural role and the behavior of “living active ingredients” for beneficial organisms.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Addison JA (1993) Persistence and nontarget effects of Bacillus thuringiensis in soil: a review. Can J For Res 23(11):2329–2342. https://doi.org/10.1139/x93-287
Alaux C, Ducloz F, Crauser D, Le Conte Y (2010) Diet effect on honeybee immunocompetence. Biol Lett 6:562–565. https://doi.org/10.1098/rsbl.2009.0986
Bailey J, Scott-Dupree C, Harris R, Tolman J, Harris B (2005) Contact and oral toxicity to honey bees (Apis mellifera) of agents registered for use for sweet corn insect control in Ontario, Canada. Apidologie 36(4):623–633. https://doi.org/10.1051/apido:2005048
Bravo A, Gill SS, Soberòn M (2007) Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 49:423–435. https://doi.org/10.1016/j.toxicon.2006.11.022
Bravo A, Likitvivatanavong S, Gill SS, Soberón M (2011) Bacillus thuringiensis: a story of a successful bioinsecticide. Insect Biochem Mol Biol 41(7):423–431
Brighenti DM, Carvalho CF, Carvalho GA, Brighenti CRG, Carvalho SM (2007) Bioatividade do Bacillus thuringiensis var. kurstaki (Berliner, 1915) para adultos de Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae). Ciência e Agrotecnologia, Lavras 31(2):279–289. https://doi.org/10.1590/S1413-70542007000200003
D’Urso V, Mazzeo G, Vaccalluzzo V, Sabella G, Bucchieri F, Viscuro R, Vitale DGM (2017) Observations on midgut of Apis mellifera workers (Hymenoptera: Apoidea) under controlled acute exposures to a Bacillus thuringiensis based biopesticide. Apidologie, Paris 48(1):5162. https://doi.org/10.5433/1679-0359.2018v39n1p329
Davidson G, Phelps K, Sunderland KD, Pell JK, Ball BV, Shaw KW, Chandler D (2003) Study of temperature growth interactions of entomopathogenic fungi with potential for control of Varroa destructor (Acari: Mesostigmata) using a nonlinear model of poikilotherm development. J Appl Microbiol 94:816–825. https://doi.org/10.1046/j.1365-2672.2003.01871.x
Davis AR (1989) Study of insecticide poisoning of honey bee brood. Bee World 70:163–174. https://doi.org/10.1080/0005772x.1989.11099013
DeGrandi-Hoffman G, Chen Y (2015) Nutrition, immunity and viral infections in honey bees. Curr Opin Insect Sci. 10:170–176. https://doi.org/10.1016/j.cois.2015.05.007
DeGrandi-Hoffman G, Chen Y, Huang E, Huang MH (2010) The effect of diet on protein concentration, hypopharyngeal gland development and virus load in worker honey bees (Apis mellifera L.). J Insect Physiol 56(9):1184–1191. https://doi.org/10.1016/j.jinsphys.2010.03.017
Dietrich S, Scholz-Döbelin H, Reintges T, Pelz J, Jehle JA, Keßler J (2014) Investigations on the residues of XenTari® (Bacillus thuringiensis subspec. aizawai) on greenhouse tomatoes. Journal für Kulturpflanzen. 66(9):312–318. https://doi.org/10.5073/JFK.2014.09.04
Duan JJ, Marvier M, Huesing J, Dievly G, Huang ZY (2008) A meta-analysis of effects of Bt crops on honey bees (Hymenoptera: Apidae). PLoS ONE 3(1):e1415. https://doi.org/10.1371/journal.pone.0001415
Engel P, Kwong WK, McFrederick Q, Anderson KE, Barribeau SM, Chandler JA, Cornman RS et al (2016) The bee microbiome: impact on bee health and model for evolution and ecology of host-microbe interactions. mBio 7(2):e02164-15. https://doi.org/10.1128/mBio.02164-15
Feitelson JS (1993) The Bacillus thuringiensis family tree. In: Kim L (ed) Advanced engineered pesticides. Marcel Dekker, Inc., New York, pp 63–67
Feitelson JS, Payne J, Kim L (1992) Bacillus thuringiensis: insects and beyond. Bio/Technology 10:271–275. https://doi.org/10.1038/nbt0392-271
Haddad ML, Polanczyk RA, Alves SB, Garcia MO (2005) Field persistence of Bacillus thuringiensis on maize leaves (Zea mays L.). Braz J Microbiol 36:309–314. https://doi.org/10.1590/S1517-83822005000400001
Ignoffo CM, Garcia C (1978) UV-photoinactivation of cells and spores of Bacillus thuringiensis and effects of peroxidase on inactivation. Environ Entomol 7(2):270–272
Kassambara A, Kosinski M, Biecek P (2019) survminer: Drawing Survival Curves using ‘ggplot2’. Avaliable at: http://www.sthda.com/english/rpkgs/survminer/
Köhl J, Booij K, Kolnaar R, Ravensberg WJ (2019) Ecological arguments to reconsider data requirements regarding the environmental fate of microbial biocontrol agents in the registration procedure in the European Union. BioControl 64:469–487. https://doi.org/10.1007/s10526-019-09964-y
Kwong WK, Moran NA (2016) Gut microbial communities of social bees. Nat Rev Microbiol 14:374–348. https://doi.org/10.1038/nrmicro.2016.43
Li SY, Fitzpatrick S, Isman MB (1995) Effect of temperature on toxicity of Bacillus thuringiensis to the oliquebanded leafroller (Lepidoptera: Tortricidae). Can Entomol 127(2):271–273. https://doi.org/10.4039/Ent127271-2
Libardoni G, de Gouvea A, Costa-Maia FM, Lozano ER, de Freitas PF, Colombo FC, Raulino F, Maciel RMA, Potrich M (2018) Effect of different Bacillus thuringiensis strains on the longevity of Africanized honey bee. Semina: Ciências Agrárias, Londrina 39(1):329–338. https://doi.org/10.5433/1679-0359.2018v39n1p329
Malone LA, Burgess EPJ, Stefanovic D (1999) Effects of a Bacillus thuringiensis toxin, two Bacillus thuringiensis biopesticide formulations, and a soybean trypsin inhibitor on honey bee (Apis mellifera L.) survival and food consumption. Apidologie. 30:465–473
Mommaerts V, Jans K, Smagghe G (2010) Impact of Bacillus thuringiensis strains on survival, reproduction and foraging behaviour in bumble bees (Bombus terrestris). Pest Manag Sci. 66:520–525. https://doi.org/10.1002/ps.1902
Moran NA, Hansen AK, Powell JE, Sabree ZL (2012) Distinctive gut microbiota of honey bees assessed using deep sampling from individual worker bees. PLoS One 7:e36393
Motta EV, Raymann K, Moran NA (2018) Glyphosate perturbs the gut microbiota of honey bees. Proc Natl Acad Sci 115(41):10305–10310
Ogunjimi AA, Gbenle GO, Akinrimisi EO (2000) PCR-based identification of Bacillus thuringiensis isolated from soil samples in Nigeria. Zeitschrift für Naturforschung C 55(11–12):987–990
Organisation for Economic Co-operation and Development (OECD) (2016) Guidance document on honey bee larval toxicity test following repeated exposure. Series on Testing & Assessment No. 239. ENV/JM/MONO (2016) 34. https://one.oecd.org/document/ENV/JM/MONO(2016)34/en/pdf
Organisation for Economic Co-operation and Development (OECD) (2017) OECD Guideline for the Testing of Chemicals. Honey bee (Apis mellifera), chronic oral toxicity test (10-day feeding). OECD/OCDE 245. Adopted 9 October 2017. https://www.oecd-ilibrary.org/docserver/9789264284081-en.pdf?expires=1586028842&id=id&accname=guest&checksum=F78F26502CCEE0CBABA58FF0475031F4
Pardo-López L, Soberón M, Bravo A (2013) Bacillus thuringiensis insecticidal three-domain Cry toxins: mode of action, insect resistance and consequences for crop protection. FEMS Microbiol Rev. 37:3–22. https://doi.org/10.1111/j.1574-6976.2012.00341.x
Pedersen JC, Damgaard PH, Eilenberg J, Hansen BM (1995) Dispersal of Bacillus thuringiensis var. kurstaki in an experimental cabbage field. Can J Microbiol. 41:118–125
Pinnock DE, Brand RJ, Milstead JE (1971) Field persistence of Bacillus thuingiensis spores. J Invert Pathol 18(3):405. https://doi.org/10.1590/S1517-83822005000400001
Potrich M, da Silva RT, Maia F, Lozano ER, Rossi RM, Colombo FC, Tedesco FG, Gouvea AD (2018) Effect of entomopathogens on Africanized Apis mellifera L. (Hymenoptera: Apidae). Revista Brasileira de Entomologia 62(1):23–28. https://doi.org/10.1016/j.rbe.2017.12.002
R Core Team (2019) R: A language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria
Renzi MT, Amichot M, Pauron D, Tchamitchian S, Brunet JL, Kretzschmar A, Maini S, Belzunces LP (2016) Chronic toxicity and physiological changes induced in the honey bee by the exposure to fipronil and Bacillus thuringiensis spores alone or combined. Ecotoxicol Environ Saf 127:205–213. https://doi.org/10.1016/j.ecoenv.2016.01.028
Schnepf E, Crickmore N, Van Rie J, Lereclus D, Baum J, Feitelson J, Zeigler DR, Dean DG (1998) Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol Mol Biol Rev 62:775–806
Schünemann R, Knaak N, Fiuza LM (2014) Mode of action and specificity of Bacillus thuringiensis toxins in the control of caterpillars and stink bugs in soybean culture. ISRN Microbiol 12 p. https://doi.org/10.1155/2014/135675
Soni J, Thakur M (2011) Effect of biopathogens on honey bees. Pest Technology. Glob Sci Books 5(1):86–90
Southwick EE, Heldmaier G (1987) Temperature control in honey bee colonies. Bioscience. 37:395–399. https://doi.org/10.2307/1310562
Vandenberg JD, Shimanuki H (1986) Two commercial preparations of the β exotoxin of Bacillus thuringiensis influence the mortality of caged adult honey bees, Apis mellifera (Hymenoptera: Apidae). Environ Entomol 15(1):166–169. https://doi.org/10.1093/ee/15.1.166
Vandenberg JD, Shimanuki H (1990) Viability of Bacillus thuringiensis and its efficacy for larvae of the greater wax moth (Lepidoptera: Pyralidae) following storage of treated combs. J Econ Entomol 83:760–765
Winston ML (1987) The biology of the honey bee. Harvard University Press
Acknowledgments
The authors are grateful to the employees of the research apiary at the JKI Braunschweig for their technical assistance. We thank the team of the Institute of Geoecology at the Technische Universität Braunschweig for helpful discussions.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
CS and ATA idealized the work and performed the statistical analysis of the collected data. JHE assisted in the conduction of the larval experiment. HB performed the analysis of laboratory procedures. JP and DR supervised the work. All authors participated in the writing and reviewing the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Responsible editor: Giovanni Benelli
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Steinigeweg, C., Alkassab, A.T., Beims, H. et al. Assessment of the impacts of microbial plant protection products containing Bacillus thuringiensis on the survival of adults and larvae of the honeybee (Apis mellifera). Environ Sci Pollut Res 28, 29773–29780 (2021). https://doi.org/10.1007/s11356-021-12446-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-021-12446-3