
Abstract
Carassius carp is one of the most popular freshwater fish as a source of food and aesthetic enjoyment from historical times. However, the species status of this group is controversial owing to extreme morphological variation and various ploidy levels, and therefore, many regional groups still remain to be reconsidered. The taxonomy of the Carassius fish in the Korean Peninsula, previously identified as C. auratus langsdorfi, has long been in doubt because several river basins were isolated from each other by the sea to such an extent that the strong geographic structure of the population may have influenced this fish group distributed over whole peninsula. Here, we suggest some clues to resolve the taxonomic entity of Carassius fish in South Korea using mitochondrial genetic variation in a comparison with those from nearby regions with a view to establishing the phylogeography and genetic structure of population. We found three monophyletic phylogroups (pgA, pgB, and pgC), which interestingly do not relate to C. auratus langsdorfi. Geographically, pgA and pgB were observed only in the westward rivers (WWRs) and southward rivers (SWRs), respectively, except for some marginal locations between the two river groups. pgA, including the haplotypes from Weihai, northern China, shared a common ancestry with C. gibelio. pgC may be a lineage that has been introduced, as shown by the result that it belongs to same clade as goldfish. It is likely that pgB has undergone a distinct evolutionary process after separation from pgA and the population of northern mainland China. Our results suggest the need of further taxonomic studies to elucidate the status of these groups and to assign an adequate taxonomic category to each. And, therefore, pgA and pgB, regardless of taxonomic level, deserve conservation as endemic groups in this region.
Similar content being viewed by others

Introduction
Carassius carp is one of the most popular freshwater fish groups that mankind has been concerned with. It has been a valuable food source from ancient times, and has been considered as an attraction encouraging recreational fishing activities (Jung et al., 2007). In addition, developed from colored variants of Carassius, the goldfish, Carassius auratus auratus (Linnaeus, 1758) has been the most popular aquarium fish. For these reasons, this fish group now has worldwide distribution (Courtenay et al., 1974). It was frequently reported that Carassius disturbed freshwater ecosystems in introduced regions (Fraser & Adams, 1997), which makes this fish group ecologically remarkable, too.
According to Species 2000 & ITIS catalogue of life 2009 (Bisby et al., 2009), the genus Carassius is composed of four species: Carassius carassius (Linnaeus, 1758) (Crucian carp), C. cuvieri Temminck & Schlegel, 1846 (gengorobuna), C. gibelio (Bloch, 1782) (Prussian carp), and C. auratus (Linnaeus, 1758). And the latter species, dependent on its geographic distribution, is divided into five recorded subspecies—C. a. auratus (Linnaeus, 1758) (goldfish), C. a. argenteaphthalmus Nguyen, 2001, C. a. langsdorfi Temminck & Schlegel, 1846 (ginbuna), C. a. grandoculis Temminck & Schlegel, 1846 (nigorobuna), and C. a. buergeri Temminck & Schlegel, 1846 (okinbuna)—and a lot of unnamed regional variants which remain to be determined. However, species status of belonging to this genus still remains controversial (Iguchi et al., 2003). Taxonomic ascertainment and identification of this group is not easy owing to the existence of multiple reproductive modes such as hybridogenesis and gynogenesis primarily caused by complicated ploidy level (Murakami & Fujitani, 1997; Zhou et al., 2000; Brykov et al., 2002; Zhou & Gui, 2002; Iguchi et al., 2003) and the possibility of hybridization with closely related species (Ohara et al., 2000; Hanfling et al., 2005; Toth et al., 2005) as well as extremely high morphological variation. Therefore, molecular approaches have been preferred to circumvent such difficulties, and many of referred matters, as a result, could have been solved.
Located between mainland China (or Asia) and the Japanese Islands, the fish fauna of the Korean Peninsula must have been influenced by nearby regions showing discrete features. In his monographic study on freshwater fish, Kim (1997) suggested the demarcation of rivers in the Korean Peninsula into three biogeographic subdistricts based on distribution of freshwater fish species. Each subdistrict consists of rivers isolated from others by sea but related to adjacent regions outside the Peninsula such as mainland China and the Japanese Islands through connected rivers during the glacial periods in the Quaternary. Occurring in all three biogeographic districts, Carassius would be a good material for biological questions such as whether its population is genetically structured across biogeographic subdistricts, if so then how much divergence is there across subdistricts, and finally, what taxonomic entity is. The taxonomic status of Korean Carassius fish is not clear. Although there had been some disputes, there has been a well-established agreement on Carassius fish in South Korea. Yang (1985) examined allozyme variations and morphological traits of seven Carassius populations in South Korea and compared them with those of several subspecies of Carassius auratus from the Japanese Islands. He concluded that the Korean population of Carassius was composed of C. a. langsdorfi, and that no significant divergence was detected among the populations he examined, although there was considerable morphological and molecular diversity within the populations. This conclusion was supported by morphological studies (e.g., Nam et al., 1989). However, as his conclusions were reached through comparison only with Japanese subspecies available at that time, his decision may not be persuasive since the fish populations of mainland China as well as those of the Japanese Islands shared biogeographic processes with the Korean Peninsula during the Pleistocene, and so must be closely related with each other. In addition, the protein electrophoresis method he used does not generally ensure good resolution for precise population genetic studies Therefore, the taxonomic status of Korean Carassius needs to be reconsidered.
In this study, we analyzed the genetic structure of populations of Carassius fish from most of the river systems in South Korea using mitochondrial control region sequences and compared them with those of nearby regions outside the Korean Peninsula.
Materials and methods
Sampling and DNA extraction
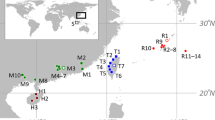
South Korea, situated in the southern part of the Korean Peninsula, has a number of rivers, most of which belong to the basins of the five major rivers, namely, the Han, Nakdong, Geum, Seomjin, and Yeongsan rivers. The majority of them (The Han, Geum, and Yeongsan) flow into the Yellow Sea, the sea to the west of Korea, and the others (The Nakdong and Seomjin) flow into the South Sea (Fig. 1). We collected individual specimens of Carassius from 18 locations in South Korea, including the basins of the five major rivers (Fig. 1; Table 1) using fishing nets or traps. The fish samples from Weihai (Shantung Province, China) were bought from the fish market. The samples were fixed using absolute ethanol immediately after capture. A total genomic DNA extraction was performed using the standard phenol extraction procedure (Sambrook & Russell, 2001).

A map denoting the sampling locations in this study and the major rivers in South Korea
PCR and sequencing
Polymerase chain reactions (PCRs) were performed using the primers designed by our team to amplify the mitochondrial control region (mtCONF: 5′-TTY TAA CTC WCA CCC CTR RCT MCC-3′, and mtCONR: 5′-GTK GCT GKC ACG AGT TTT ACC GGC-3′). The reaction mixtures were composed of 1× Taq polymerase buffer, 1 mmol/l dNTP mix, 2.5 mmol/l MgCl2, 0.5 μmol/l of each primer, 2.5–250 pg total DNA, and 0.04 U/μl Taq DNA polymerase (Promega, Madison, WI). PCR was performed on a GeneAmp 9700 unit (Applied Biosystems, Foster City, CA) and included the following steps: initial denaturation at 94°C for 3 min, 35 cycles of denaturation at 94°C for 30 s, annealing at 55°C for 40 s, elongation at 72°C for 1 min, and final extension at 72°C for 1 min. The amplified products from the PCR mixtures were purified using a QIAquick PCR purification kit (Qiagen, Valencia, CA). Subsequently, the PCR products were sequenced using both the amplifying primers and the Big Dye terminator sequencing kit (Applied Biosystems) according to the manufacturer’s instructions. The purified products were then analyzed on a Genetic Analyzer 3730 (Applied Biosystems).
Data analysis
We determined the sequences from 126 individuals of Carassius fish; thereafter, the sequences were aligned using ClustalX (Thompson et al., 1997) and exported to the NEXUS and PHYLIP formats for data analyses. DnaSP 4.10.9 (Rojas et al., 2003) was used to define the haplotypes from the aligned data sets. We used ARLEQUIN 3.11 (Excoffier et al., 2005) to calculate the gene and nucleotide diversity and the scaled mutation rate, theta estimates based on the average number of pairwise differences (pi), and the number of segregating sites (S). Using MEGA4 (build #4025) (Tamura et al., 2007) along with the neighbor-joining and maximum parsimony algorithms, the phylogenetic relationships among the haplotypes in this study and among the sequences retrieved from GenBank: C. a. langsdorfi (AB006953) (Murakami et al., 1998), C. a. auratus (AB111951), C. gibelio (EF633632 and EF633680), and C. cuvieri (AB045144), were determined. The maximum composite likelihood was chosen as the model for sequence evolution in performing neighbor-joining analysis. The average p-distances between the phylogroups were calculated using MEGA4. Computation of the genetic distances between the subpopulations was performed using ARLEQUIN 3.11, and the corrected pairwise differences calculated by subtracting the mean of the difference within the populations from the mean number of pairwise differences between the populations. The genetic relationships between the local populations were schematized by the neighbor-joining clustering method based on the matrix of corrected average pairwise differences using MEGA4. Mismatch distribution analysis (Harpending, 1994) and Tajima’s (1989) and Fu’s (1997) neutrality tests were conducted using ARLEQUIN 3.11 to infer the recent demographic history.
Results
Haplotypes and genetic diversity
A total of 49 haplotypes (GenBank accession numbers, FJ817294–FJ817342) (1,056 bp) were identified among the 126 individual specimens of Carassius from Korea and China. Three monophyletic groups—Phylogroups A, B, and C—could be observed by both of the neighbor-joining and maximum parsimony methods. As both show nearly same topology, the latter is not shown in Fig. 2. Phylogroup A (pgA), Phylogroup B (pgB), and Phylogroup C (pgC) were found to be composed of 31 haplotypes (64 individuals), 10 haplotypes (28 individuals), and 8 haplotypes (34 individuals), respectively (Table 1). pgA and pgB were more closely related to each other than to pgC. Interestingly, the haplotypes from Weihai, China, belonged to pgA together with the sequences of C. gibelio (EF633632 and EF633680), and goldfish (C. a. auratus) was included in pgC. C. a. langsdorfi, to our surprise, did not correspond closely to any phylogroup from South Korea, and its sequence showed a relatively large divergence from both pgA and pgB (4.4% of the p-distance) as compared to that between C. gibelio and pgA (0.5%) or pgB (1.6%). This result does not conform to the previous conclusion that Carassius fish in the Korean Peninsula were C. a. langsdorfi (Yang, 1985; Nam et al., 1989). C. cuvieri, an endemic species from the Japanese Islands, showed the largest divergence from pgA and pgB (average, 5.6%; Fig. 2). When the locations in South Korea were compared, the differences between the gene diversity and nucleotide diversity of locations may have arisen from the fact that locations showing higher nucleotide diversity generally have heterogeneous haplotype compositions from several phylogroups (Table 1).

A neighbor-joining tree depicting the genetic relationship among the haplotypes in this study and the sequences from several subspecies or species belonging to the genus Carassius. The geographic origin of the sequences and their GenBank accession numbers are indicated in parentheses and brackets, respectively. The numbers on the nodes represent the bootstrapping confidence values (neighbor-joining method/maximum parsimony method)
Geographical distribution of haplotypes
In the geographical distributional pattern, the haplotypes belonging to pgA were dominant in the westward rivers (WWRs) such as The Han, Geum, and Yeongsan, while those belonging to pgB comprised the majority in the southward rivers (SWRs) such as The Nakdong and Seomjin (Fig. 3). The haplotypes of pgA and pgB coexisted in Yangang, Euiseong, and Woopo, which are located at the boundary between the SWRs and WWRs. The haplotypes of pgC were distributed over both the river groups and did not show any discrete pattern of distribution as pgA and pgB.

Geographic distribution of the three phylogroups—pgA, pgB, and pgC—in South Korea. The proportions of individuals belonging to pgA (black), pgB (gray), and pgC (white) are represented in the pie chart. The dotted line denotes the border between the westward rivers (WWRs) and the southward rivers (SWRs)
The locations of all SWRs and WWRs except Woopo formed monophyletic groups in the neighbor-joining tree, as inferred from the distance matrix of the corrected pairwise differences (Fig. 4). Despite the fact that Woopo belonged to the SWRs, it was included into the WWR clade in the population tree. This may have arisen from the fact that most of its sequences were from pgA and pgB rather than from pgB alone (Fig. 3; Table 1). Euiseong and Yangang were located between the SWRs and WWRs in the population tree, which may have been due to the lineage admixture of pgA and pgB.

A neighbor-joining phenogram denoting the genetic relationship among the demes in this study
Genetic properties and the demographic history of phylogroups
When the genetic properties of the phylogroups were compared, pgA was shown to exhibit increases in most of the genetic diversity indices compared to the other two groups (Table 2). All the estimates of pgB were slightly lower than those of pgA, but those of pgC were the smallest in all the summary statistics for genetic diversity. pgA and pgB showed a considerable discrepancy in theta (S), but little difference in theta (pi). The theta estimates of pgC were also fairly lower than those of the others. Statistically significant negative values of Tajima’s D and Fu’s Fs were observed in pgA and pgC, which implies that these groups underwent recent population expansion under the assumption of selective neutrality. However, the corresponding values of pgB were not statistically significant. This observation was supported by mismatch distribution analyses where the observed patterns of pgA and pgC conformed well to those simulated under the model of sudden expansion of population, but pgB presented considerable differences (Fig. 5). Thus, the above results imply that pgB may have undergone evolutionary processes different from those that pgA and pgC underwent.

The observed mismatch distributions (open bars) of phylogroups A, B, and C with the expected distributions (filled bars) simulated under the sudden expansion model of population fitted to the observed distributions
Discussion
Owing to the discrete geographic patterns, it can be inferred that the population genetic structure of the Carassius fish in the Korean Peninsula may have been significantly influenced by geological processes in the past. It is known that geological changes occurring during the late Cenozoic era have had a great influence on the biogeography of fish fauna (Bernatchez & Wilson, 1998). The topography near the Korean Peninsula has undergone a great change due to the climatic changes during the Quaternary (Chough, 1983). Owing to the nearby extensive continental shelf, the freshwater environment of the northern Asian region was favorable for the evolution of speciose fish fauna during this period because of the connections formed between the streams and rivers (dispersal) with the lowering of the sea level, followed by disconnection (vicariance) owing to the raised sea level (Kim, 1997). The previous connections between the rivers in mainland China and those of the WWRs in Korea could be evidenced from the result that the haplotypes from Weihai, China, and the WWRs of South Korea shared a recent common ancestry.
Not all the rivers in the Korean Peninsula, however, showed a previous connection with those of mainland China. The SWRs of the Korean Peninsula did not show geologically close relationships with the WWRs because the former may have been disconnected from the rivers of northern China earlier than the latter. pgB mostly occurred in the SWRs forming reciprocal monophyly without containing any haplotypes from China. The relatedness between mainland China and the WWRs and the relative independence of the SWRs from the WWRs were suggested in a study of medaka fish from the Korean Peninsula and China (Takehana et al., 2004). Further, this inference is also supported by the fact that many of the species (42 species) do not coexist across the WWRs and SWRs, as shown in the study of the biogeography of 93 cyprinids and silurids fish species in Korea (Kim, 1997). In the same study, the speciation of fish through vicariance across the two river groups of the Korean Peninsula was proven by considering the geographic distribution of the six sister species of Iksookimia endemic to that region. Therefore, the biological diversity of freshwater fish in South Korea could be greatly attributed to the divergence of the population and to the speciation processes across these two river groups caused by geological changes during the late Cenozoic era. Kim (1997) suggested the demarcation of rivers of the Korean Peninsula into three biogeographic subdistricts: the West Korea subdistrict, which was the same as the WWRs in our study; the South Korea subdistrict, which was the same as the SWRs; and the East–North Korea subdistrict, including the rivers located in the northeast portion of the Korean Peninsula.
The co-occurrence of haplotypes from pgA and pgB at some locations situated in the margin between the WWRs and SWRs may be partly due to river capture (Bishop, 1995). Most of the major rivers of South Korea generally originate in the inland mountains, and their origins are, therefore, situated close to each other. Mixing of the waters in the upper streams and the resulting admixture of haplotypes across geographic barriers, therefore, may occur due to topographic changes near the river origins. This scenario is highly possible given the fact that a few of the endemic Korean species are commonly distributed over the WWRs and SWRs (Kim, 1997). Intentional or unintentional introduction across rivers may partly explain the admixture pattern of pgA and pgB. Carassius fish are such popular fish for fishing and food that some individuals may have been introduced into the rivers other than those that originate from fish rearing and the releasing ceremonies performed by the believers of Buddhism.
Phylogroup C (pgC) appears to be a lineage that has recently been introduced into South Korea. It does not represent the significant differences in the geographic structures across the rivers as pgA and pgB do; further, it shows low genetic diversity, which is typical of introduced populations. pgC may be related to goldfish, which can be inferred from the fact that they form a monophyletic group. This guess, if true, could be explained by the naturalization of goldfish into the freshwater ecosystem of the Korean Peninsula. Goldfish were introduced so early in the sixteenth century that it is the first introduced fish species in the world (Lorenzoni et al., 2007). Some of them were naturalized in non-native environments and have disturbed the freshwater ecosystems of the region in which they were introduced (Vekhov, 2008). They could threaten the endemic fish fauna not only by competing with the other fish species occupying the same ecological niche but also by disturbing the genetic identity of the endemic fish fauna through hybridization (reviewed in Lorenzoni et al., 2007). Careful observations should be performed in the near future to assess the effect of the introduced lineage on the freshwater ecosystem of the Korean Peninsula.
From the phylogeographic view, our results showed that pgA and pgB exhibit a closer relationship with those from mainland China rather than those from the Japanese Islands including C. a. langsdorfi, a previously identified species in this country (Fig. 2). These are consistent with previous studies on other fish species in which those from the Korean Peninsula are more closely related to those of mainland China than with those of the Japanese Islands (Okazaki et al., 2002; Takehana et al., 2004). Although our study was performed using only one locus, that is, the mitochondrial control region, it provides sufficient clues about the species entity and taxonomic composition of Carassius. However, further detailed taxonomic studies are required on the phylogroups of Carassius fish identified in this study to know the exact taxonomic entity and assign them adequate categorical levels. And these studies should be performed based on a diverse range of evidence, including morphometric data, ploidy, and molecular genetic variation.
Both pgA and pgB satisfy the criteria of evolutionarily significant units (ESUs) suggested by Crandall et al. (2000), that is, reciprocal monophyly and discrete geographic distribution; therefore, regardless of their taxonomic status, they deserve conservation. Ambitious projects to repair waterways and construct huge inland water canals, which would connect the WWRs and the SWRs, may be initiated in the near future in South Korea. However, such projects are expected to not only cause severe destruction of the habitats of most freshwater species, including Carassius, but also inhibit evolutionary processes, such as the divergence of population across isolated rivers from which the biological diversity of freshwater ecosystems has arisen. Hence, there is an urgent requirement for a measurement system to conserve the identity of the population inhabiting the freshwater ecosystems.
References
Bernatchez, L. & C. C. Wilson, 1998. Comparative phylogeography of Nearactic and Palearctic fishes. Molecular Ecology 7: 431–452.
Bisby, F. A., Y. R. Roskov, T. M. Orrell, D. Nicolson, L. E. Paglinawan, N. Bailly, P. M. Kirk, T. Bourgoin & G. Baillargeon (eds), 2009. Species 2000 & ITIS Catalogue of Life: 2009 Annual Checklist. Digital resource at www.catalogueoflife.org/annual-checklist/2009/. Species 2000, Reading, UK.
Bishop, P., 1995. Drainage rearrangement by river capture, beheading and diversion. Progress in Physical Geography 19: 449–473.
Brykov, Vl. A., N. E. Polyakova, L. A. Skurikhina, S. M. Dolganov, M. G. Eliseikina & M. Yu. Kovalev, 2002. Mitochondrial DNA variation in goldfish Carassius auratus gibelio from Far Eastern water reservoirs. Russian Journal of Genetics 38: 1176–1180.
Chough, S. K., 1983. Marine Geology of Korean Seas. International Human Resources Development Corporation, Boston.
Courtenay, W. R., J. F. Sahlman, W. W. Miley & D. J. Herrema, 1974. Exotic fishes in fresh and brackish waters of Florida. Biological Conservation 6: 292–302.
Crandall, K. A., O. R. P. Bininda-Emonds, G. M. Mace & R. K. Wayne, 2000. Considering evolutionary processes in conservation biology. Trends in Ecology and Evolution 15: 290–295.
Excoffier, L., G. Laval & S. Schneider, 2005. Arlequin ver 3.0: an integrated software package for population genetics data analysis. Evolutionary Bioinformatics Online 1: 47–50.
Fraser, D. & C. E. Adams, 1997. A crucian carp Carassius carassius (L.) in Loch Rannoch, Scotland: further evidence of the threat posed to unique fish communities by introduction of alien fish species. Aquatic Conservation 7: 323–326.
Fu, Y. X., 1997. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 147: 915–925.
Hanfling, B., P. Bolton, M. Harley & R. Carvalho, 2005. A molecular approach to detect hybridisation between crucian carp (Carassius carassius) and non-indigenous carp species (Carassius spp. and Cyprinus carpio). Freshwater Biology 50: 403–417.
Harpending, R. C., 1994. Signature of ancient population growth in a low-resolution mitochondrial DNA mismatch distribution. Human Biology 66: 591–600.
Iguchi, K., G. Yamamoto, N. Matsubara & M. Nishida, 2003. Morphological and genetic analysis of fish of a Carassius complex (Cyprinidae) in Lake Kasumigaura with reference to the taxonomic status of two all-female triploid morphs. Biological Journal of Linnean Society 79: 351–357.
Jung, J., E. Lee & W. Kim, 2007. Isolation and characterization of polymorphic trinucleotide microsatellites of the polyploid crucian carp (Carassius auratus). Molecular Ecology Notes 7: 124–126.
Kim, I. S., 1997. Illustrated Encyclopedia of Fauna and Flora of Korea. Vol. 37, Freshwater Fishes. Ministry of Education of Korea, Seoul (in Korean).
Lorenzoni, M., M. Corboli, L. Ghetti, G. Pedicillo & A. Carosi, 2007. Growth and reproduction of the goldfish Carassius auratus: a case study from Italy. In Gherardi, F. (ed.), Biological Invaders in Inland Waters: Profiles, Distribution and Threats. Springer, Dordrecht, The Netherlands: 259–273.
Murakami, M. & H. Fujitani, 1997. Polyploid-specific repetitive DNA sequences from triploid ginbuna (Japanese silver crucian carp, Carassius auratus langsdorfi). Genes & Genetic Systems 72: 107–113.
Murakami, M., Y. Yamashita & H. Fujitani, 1998. The complete sequence of mitochondrial genome from a gynogenetic triploid “Ginbuna” (Carassius auratus langsdorfi). Zoological Science 15: 335–337.
Nam, M. M., H. J. Yang & B. K. Seo, 1989. Morphological variation of the crucian carp, Carassius auratus (Linnaeus) from Yongnam area in Korea. Korean Journal of Ichthyology 1: 54–63.
Ohara, K., T. Ariyoshi, E. Sumida, K. Sitizyo & N. Taniguchi, 2000. Natural hybridization between diploid crucian carp species and genetic independence of triploid crucian carp elucidated by DNA markers. Zoological Science 17: 357–364.
Okazaki, T., S. R. Jeon & T. Kitagawa, 2002. Genetic differentiation of piscivorous chub (genus Opsariichthys) in Japan, Korea and Russia. Zoological Science 19: 601–619.
Rojas, J., J. C. Sanchez-DelBarrio, L. Messeguer & R. Rozas, 2003. DnaSP, DNA polymorphisms analyses by the coalescent and other methods. Bioinformatics 19: 2496–2497.
Sambrook, J. & D. W. Russell, 2001. Molecular Cloning: A Laboratory Manual, 3rd ed. Cold Spring Harbor Press, Cold Spring Harbor.
Tajima, F., 1989. The effect of change in population size on DNA polymorphism. Genetics 123: 597–601.
Takehana, Y., S. Uchiyama, M. Matsuda, S. R. Jeon & M. Sakaizumi, 2004. Geographic variation and diversity of cytochrome b gene in wild populations of Medaka (Oryzias latipes) from Korea and China. Zoological Science 21: 483–491.
Tamura, K., J. Dudley, M. Nei & S. Kumar, 2007. MEGA 4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Molecular Biology and Evolution 24: 1596–1599.
Thompson, J. D., T. J. Gibson, F. Plewniak & D. G. Higgins, 1997. The Clustal X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research 24: 4876–4882.
Toth, B., E. Varkonyi, A. Hidas, E. E. Meleg & L. Varadi, 2005. Genetic analysis of offspring from intra- and interspecific crosses of Carassius auratus gibelio by chromosome and RAPD analysis. Journal of Fish Biology 66: 784–797.
Vekhov, D. A., 2008. Population of silver crucian carp Carassius auratus (Cypriniformes, Cyprinidae) with “Golden” individuals in the pond of the city of Volgograd. Journal of Ichthyology 48: 326–335.
Yang, S. Y., 1985. A study on the systematic status Korean crucian carp (Carassius auratus). Bulletin of the Institute for Basic Science Inha University 6: 155–161 (in Korean).
Zhou, L. & J. F. Gui, 2002. Karyotytic diversity in polyploid gibel carp, Carassius auratus gibelio Bloch. Genetica 115: 223–232.
Zhou, L., Y. Wang & J. F. Gui, 2000. Genetic evidence for gonochoristic reproduction in gynogenetic silver crucian carp (Carassius auratus gibelio Bloch) as revealed by RAPD assays. Journal of Molecular Evolution 51: 498–506.
Acknowledgments
This study was supported by the Brain Korea 21 project of Ministry of Education, and by the Korea Research Foundation Grant funded by the Korean Government (MOEHRD) (KRF-2005-2008-314-C00379).
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: C. Sturmbauer
Rights and permissions
About this article
Cite this article
Jung, J., Song, K.H., Lee, E. et al. Mitochondrial genetic variations in fishes of the genus Carassius from South Korea: proximity to northern China rather than Japanese Islands?. Hydrobiologia 635, 95–105 (2009). https://doi.org/10.1007/s10750-009-9866-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-009-9866-x