
Abstract
Agroforestry is a common traditional practice in China, especially in Southern Xinjiang, Northwest China. However, the productivity of many agroforestry systems has been lower than expected in recent years. We chose an apricot (Prunus armeniaca L.)/wheat (Triticum aestivum L.) intercropping system to investigate the influence of apricot tree shade intensity on the grain yield and quality of intercropping wheat in Southern Xinjiang Province. We found that the mean daily light intensity in the east-row, inter-row, and west-row positions (apricot trees were planted in north–south orientation) in the intercropping systems decreased by 61.5%, 42.2%, and 63.6%, respectively, compared to the monoculture wheat. Correspondingly, the mean daily photosynthesis rate decreased by 52.0%, 28.4%, and 38.8%, respectively. The total florets, fertile florets, total spikelets, fertile spikelets, and grain yield and its components (including spike number, grains per spike, and thousand grain weight), and the total N and P contents of intercropping wheat in apricot-based intercropping systems were all significantly decreased compared to monoculture wheat. However, the protein content and wet gluten content of intercropped wheat were significantly increased compared to monoculture wheat. We did not find significant differences in any parameters among east-row, inter-row, and west-row positions in intercropping systems.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Agroforestry is an effective approach for land use, because the net effect of interactions between woody and herbaceous components is typically positive over the long term. Agroforestry is often promoted because it provides a number of environmental services including enhancing soil nutrient, reducing nutrient leaching, improving soil erosion, increasing soil organic carbon, and affecting the soil microbial community (Evers et al. 2010; Bergeron et al. 2011; Rivest et al. 2013; Alam et al. 2014). Intercropping of trees with crops has been practiced in China for centuries and is an important method for ensuring food security (Yang et al. 2009; Hong et al. 2017). For example, in the Loess Plateau region, young fruit trees and annual crops are a widely used agroforestry system for reducing soil erosion and water loss, and increasing economic benefits (Gao et al. 2013). In the North China Plain, Paulownia trees are usually intercropped with wheat or bean species to enhance the suitability of the microclimate (Zou and Sanford 1990).
In most tree-based intercropping systems, competition for light between trees and crops is the main reason which caused the reduction in crop yield (Artru et al. 2017). Lack of light is a major constraint that has affected stability of the structure and function of agricultural ecosystems. Kittur et al. (2016) found that low understory photosynthetically active radiation (PAR) is the dominant factor reducing growth of turmeric in dense, compared to widely spaced, bamboo stands. In apple-based intercropping systems, the total aboveground biomass and yield of peanut and soybean had highly linear correlations with PAR (Gao et al. 2013). In Xinjiang Province, the understory seed cotton yield under almond trees decreased by 66.9%, compared to monoculture cotton (Yang et al. 2009). Jose et al. (2000) observed that maize yields were reduced by 35% when alley-cropped with black walnut.
In 2012, the total area of fruit trees reached more than 1 million hectares in Southern Xinjiang, Northwest China (National Bureau of Statistics of China 2013). Intercropping systems based on fruit trees are favored by the local population because the fruit can be used to increase food security. However, the productivity of crops intercropped with fruit trees in many agroforestry systems has been lower than expected in recent years, putting pressure on the local economy (Gao et al. 2013; Zhang et al. 2017). Optimizing positive interactions while minimizing negative interactions between trees and crops requires a mechanistic understanding of belowground and aboveground interactions. In the present study, we experimentally compared growth in monocrop or an intercrop wheat (Triticum aestivum L.) under apricot trees (Prunus armeniaca L.) to examine aboveground competitive interactions and identify likely response mechanisms. Apricot tree and wheat were selected because of their importance as main economic and food crops. The objectives were to determine whether apricot trees had any significant effects on wheat growth and yield through shading, and whether apricot tree affected the quality of intercropped wheat.
Materials and methods
Experimental design
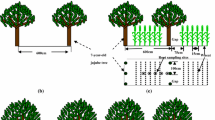
Field experiments were conducted in 2011 and 2012 in four villages of Zepu County (38°05′N, 77°10′E), Kashi Prefecture, Xinjiang Uygur Autonomous Region, China. Chemical properties of the soil are presented in Table S1. The experimental design was a field experiment, comprising monoculture wheat (T. aestivum L. Xindong-20) and wheat intercropped with 10-year-old apricot trees (P. armeniaca L. Saimaiti), and the rootstock is Armeniaca sibirica. Row distance was 0.13 m in wheat. Apricot trees were planted in north–south orientation. The basic information of apricot trees is shown in Table S2. The apricot-based intercropping wheat strip was 5.07 m wide (the wheat strip is divided equally into three regions: east-row, inter-row, and west-row positions), and the distance between the apricot tree to the nearest wheat row was 0.965 m (Fig. 1). The apricot tree occupied 27.6% of the gross area. The area of monoculture wheat and apricot tree intercropping systems was 0.4 hm2. The sown density of monoculture wheat and apricot tree-based intercropping wheat was all 4.25 × 106 plants per hm2.

Planting patterns in monoculture and the apricot–wheat-based intercropping system; the figure is schematic and does not represent the actual number of wheat rows. Er, east-row position; Ir, inter-row position; Wr, west-row position
In 2010, monoculture wheat and apricot-based intercropping wheat were all sown on October 8, 2010, and harvested on June 11, 2011, and in 2011, sown on October 3, 2011, and harvested on June 9, 2012. All fields were fertilized with farmyard manure (15,000 kg ha−1, N:P2O5:K2O = 0.37%:0.41%:0.46%), urea (275 kg N ha−1), triple superphosphate (150 kg P2O5 ha−1), and potassium sulfate (150 kg K2O ha−1). All the farmyard manure and 40% N fertilizer and all the P and K fertilizer were applied homogeneously throughout the fields before sowing wheat. The remaining 60% of the N fertilizer was applied when wheat reached the stem elongation stage. Apricot trees were not fertilized during the whole growth periods.
Harvest and analysis
Wheat was harvested when mature. In 2011 and 2012, 6.5 m2 (5.0 m length × 1.3 m width) of monoculture wheat and 6.76 m2 (4 m × 1.69 m for east-row, inter-row, and west-row positions, respectively) of wheat in an apricot-based intercropping system were harvested. There were five replicates. Samples were immediately dried on a sunny area on the ground to thresh seeds (in order to calculate wheat yield). To make wheat samples more representative, 2-m-long samples of intercropped wheat from three regions were harvested to estimate the total spike number and grains per spike, and then, all samples were threshed for seeds to estimate thousand grain weight and harvest index. The stalks (without grains) and grain samples were digested in a mixture of concentrated H2SO4 and H2O2. N concentrations were determined using the micro-Kjeldahl method and P concentrations by the molybdo-vanado-phosphate colorimetrical method (Fixen and Grove 1990). In 2011, 15 plants for each replication were selected to calculate stalk biomass at overwintering, reviving, jointing, booting, anthesis, filling, and maturity stages, respectively, and the stalk samples were heated at 105 °C for 30 min and then oven-dried (72 h, 75 °C).
Florets and spikelets
In both years, ten main spikes from each replicate, which flowered on the same day and were the same size, were destructively harvested to investigate total florets, fertile florets, total spikelets, and fertile spikelets.
Photosynthetically active radiation measurement
Light penetration was measured using a SunScan Canopy Analysis System (Delta-T Devices, Cambridge, UK), and the measured position was just above wheat (values indicate the photosynthetically active radiation (PAR) transmitted by fruit trees and incident over the wheat canopy) and underneath wheat (values indicate the PAR transmitted fruit trees and wheat canopy getting to the ground surface, TPAR) canopy. The 64 light sensors of the SunScan measured individual levels of PAR and TPAR, which are transmitted to a PDA and expressed as μmol m−2 s−1. SunScan readings were taken when the sky was clear to avoid the interference of the clouds at the filling stage of wheat in 2011. One measurement was performed every 2 h from morning at 09:00 until late afternoon at 19:00. The measurement positions were taken in the central part of each area (east-, west-, and inter-row position). An average value was calculated from two positions (the 64 light sensors were placed in north–south and east–west orientations) per replicate positioned.
Photosynthetic parameters
The net photosynthetic rate (Pn) of the flag leaves was determined with a LI-6400XT Portable Photosynthesis System (LI-COR, Inc., USA), and the readings are taken when the sky was clear to avoid the interference of the clouds at the filling stage of wheat in 2011. The measurements were conducted under traditional open system and under controlled conditions with a CO2 concentration of 380 µmol m−2 s−1. The net photosynthesis was measured under the actual PAR incident at the time of measuring. One measurement was performed every 2 h from morning at 09:00 until late afternoon at 19:00. An average value was calculated from three flag leaves per replicate positioned.
Grain quality analyses
Grain protein content (%) was determined using a near-infrared reflectance analyzer (FOSS-1241, Near Infra-Red Reflectance, Sweden), and wet gluten content was determined using Glutomatic 2200 (Perten Instruments, Huddinge, Sweden) calibrated based on official AACC methods (AACC International 2010).
Statistical analysis
One-way analysis of variance (ANOVA) was performed on all datasets using SPSS 16.0 for Windows (SPSS Inc., Chicago, IL). Significant differences between monoculture and agroforestry and within the three positions in agroforestry were determined with Duncan’s multiple range test at the 5% level. Standard error between the replicates was also calculated.
Results
Light interception
Diurnal variation of photosynthetically active radiation (PAR) and transmitted PAR (TPAR) in both the apricot-based intercropping system and the monoculture wheat had a single peak curve with time (Fig. 2). The PAR of apricot-based intercropping system was significantly lower than that in the monoculture wheat during the same period (Fig. 2a). TPAR, which indicates the PAR transmitted by fruit trees and wheat canopy and getting to the ground, was significantly lower than PAR, and all TPAR values were lower than 200 µmol m−2 s−1 (Fig. 2b). In the apricot-based intercropping system, due to the influence of the solar azimuth, the east-row position received higher PAR and TPAR in the post meridiem, whereas the west-row position correspondingly received higher PAR and TPAR in the ante meridiem.

Daily changes in photosynthetically active radiation incident over the wheat canopy (PAR) and that transmitted to the ground (TPAR) for wheat in apricot–wheat intercropping system and monoculture wheat at the filling stage in 2011. Mono, monoculture wheat; Er, east-row position; Ir, inter-row position; Wr, west-row position
Photosynthetic rate
The apricot-based intercropping wheat and the monoculture wheat showed a single peak curve with time for diurnal variation of photosynthesis rate (Pn). The Pn of monoculture wheat was higher than the intercropping wheat (Fig. 3). For example, in the intercropping system, Pn of intercropped wheat in the east-row, inter-row, and west-row positions was reduced on average by 52.0%, 28.4%, and 38.8%, respectively, compared to monoculture wheat. Similar to the findings for PAR, intercropped wheat in the east-row position had higher Pn in the post meridiem, and west-row position had higher Pn in the ante meridiem.

Daily changes in photosynthesis rate (Pn) of wheat in apricot–wheat intercropping system and monoculture wheat at the filling stage in 2011. Mono, monoculture wheat; Er, east-row position; Ir, inter-row position; Wr, west-row position
Stalk dry weight
Stalk dry weight was significantly higher in monoculture wheat than in all positions (east-row, inter-row, and west-row positions) of apricot-based intercropped wheat (Fig. 4). There were no significant differences among the three regions during the whole growth period (P < 0.05). Stalk dry weight was reduced, on average, by 21.4% for wheat in the apricot-based intercropping system compared to monoculture at the mature stage.

Stalk dry weight of wheat in apricot–wheat intercropping system and monoculture wheat at different growth stages in 2011. Mono, monoculture wheat; Er, east-row position; Ir, inter-row position; Wr, west-row position
Florets and spikelets per spike
In both years, total florets, fertile florets, total spikelets, and fertile spikelets of monoculture wheat were significantly higher than intercropped wheat (except total spikelets in 2012) (Table 1). For example, in intercropped wheat, total florets, fertile florets, total spikelets, and fertile spikelets were reduced, on average, by 25.5%, 25.8%, 4.3%, and 9.0%, respectively, in 2011, and by 26.6%, 38.2%, 4.3%, and 17.9%, respectively, in 2012 compared to monoculture wheat. There were no significant differences in total florets, fertile florets, total spikelets, or fertile spikelets of wheat in either year among east-row, inter-row, and west-row positions of intercropped wheat (Table 1).
Wheat yield components
In 2011 and 2012, the spike number, grains per spike, thousand grain weight, and net yield were significantly higher in monoculture wheat than in apricot-based intercropping wheat (except grains per spike in 2011). No significant differences were observed in either year for these characteristics among east-row, inter-row, and west-row positions (Table 2).
N and P contents
In both years, the N and P uptake of wheat in monoculture was significantly higher than west-row position in apricot-based intercropping systems (Table 3). We did not find significant differences in N and P uptake among the three positions (east-row, inter-row, and west-row positions) in 2011 and 2012 (P < 0.05).
Grain quality traits
In 2011 and 2012, the protein content and wet gluten content of wheat in apricot-based intercropping systems were significantly higher than monoculture wheat treatments (Table 4). In both years, the protein content and wet gluten content of wheat were not significantly different among east-row, inter-row, and west-row positions in the apricot-based intercropping systems.
Discussion
Photosynthetically active radiation and photosynthetic rate
Light is a primary limiting factor in agroforestry systems, and light availability highly influences the growth of intercropped crops (Chirko et al. 1996). Gao et al. (2013) observed the PAR at 0.5 m and 1.5 m distance to the tree row was reduced by 17.9% and 10.4% in apple–soybean intercropping treatment, respectively, and reduced by 17.8% and 5.4% in apple–peanut intercropping treatment. Similar results were reported for gliricidia/maize (Makumba et al. 2007), walnut/maize (Jose et al. 2000), and bamboo/turmeric (Kittur et al. 2016) studies in temperate agroforestry systems. Our results showed that the detected PAR and TPAR values of intercropped wheat were significantly lower than monoculture wheat. Considering the three inter-row positions, the PAR of east-row and west-row positions was significantly lower than the inter-row position (Fig. 2). Pn reduction depends on the shade intensity and the distance to the tree rows (Reynolds et al. 2007). For example, the understory Pn of cotton decreased by 70.2% compared to monoculture cotton in almond/cotton intercropping system in Xinjiang Province, China (Yang et al. 2009). Gao et al. (2013) observed a positive linear relationship between distance from the apple tree rows and net photosynthetic rate. The Pn of intercropped wheat was significantly influenced by the apricot tree shade (Fig. 3). Pruning and spacing could decrease shade intensity and enhance Pn of intercropped maize (Friday and Fownes 2002). Meanwhile, canopy porosity, trunk height, tree space, and fertilization significantly affect Pn of intercropped wheat as well.
Florets and spikelets
The grain number per spike is determined by the number of florets and spikelets per spike. Initiation and development of spikelets and florets in wheat are influenced by shading (Nasehzadeh and Ellis 2017; Toyota et al. 2001). Gardner et al. (1985) found a significantly negative correlation between the number of florets and spikelets with shading intensity. Our study showed that apricot tree shade significantly decreased total florets, fertile florets, total spikelets, and fertile spikelets per spike compared to monoculture wheat. No significant differences were observed for these characteristics among east-row, inter-row, and west-row positions (Table 1). Grain number per spike of wheat is affected by shading during floret growth and anthesis (Nasehzadeh and Ellis 2017), and the development of the floret varies depending on its position on the spike (Toyota et al. 2001). Willey and Holliday (1971) showed that the light intensity greatly affects spikelet initiation and floret primordia, resulting in fewer grains. For example, Toyota et al. (2001) found that shading delayed the rate of floret initiation per spike by 11.4% and consequently, decreased the number of florets by 22.3% and decreased the grain weight per spike by 19% at maturity.
Grain yield components
In general, the yield of understory crops is determined by the available light and the efficiency of intercepted light that is converted into photosynthate (Long et al. 2006; Gao et al. 2013). Light intensity influences crop photosynthetic rate, the accumulation of plant photosynthate, and yield (Rozbicki et al. 2015; Bhatta et al. 2017). The intercropped maize yields were reduced by 33% when alley-cropped with red oak (Jose et al. 2000). Zhang et al. (2017) reported that both fruit and crop yields were reduced when either 5-, 7-, or 9-year-old jujube trees were intercropped with wheat. We found that stalk biomass of intercropped wheat was highly reduced by apricot tree shading during the whole growth period, but there were no significant differences in stalk biomass among east-row, inter-row, and west-row positions (Fig. 4). However, the grain yield of durum wheat was higher in the west than in the center position in a Mediterranean alley cropping system (Inurreta-Aguirre et al. 2018). Other studies of temperate agroforestry systems found similar results: The grain yield and its components significantly decreased with decreasing the distance to the tree rows (P < 0.05) (Jaiswal et al. 1993; Peng et al. 2009; Yang et al. 2016). In the present study, grain yield and its components, including spike number, grains per spike, and thousand grain weight of intercropped wheat, were significantly decreased compared to monoculture configurations in both years (P < 0.05) (Table 2).
Stalk N and P uptake
Light plays an important role in dry matter accumulation, nutrient absorption, and yield of crops. Kittur et al. (2016) reported that the uptake of N, P, and K by understory turmeric decreased as bamboo spacing decreased. Cui et al. (2013) found that shading affects dry matter accumulation and significantly decreased the total N, P, and K contents of summer maize. In the present study, the total N and P uptake of intercropped wheat was significantly decreased in apricot-based intercropping systems; however, there were no significant differences among east-row, inter-row, and west-row positions (Table 3). It is interesting that all the differences in PAR and TPAR with the position in the inter-row did not translate into differences in morphological and physiological traits of wheat. This would indicate compensation mechanisms over the growing season for intercropped wheat. Several previous studies revealed that nutrient uptake by understory crops is strongly affected by overstory tree density, understory plant varieties, and plant nutrient demand (Rowe et al. 2001; Gao et al. 2013).
Grain quality
Light intensity is one of the most important environmental factors, influencing crop photosynthetic rate, plant growth and development, accumulation of plant photosynthate, and grain yield and quality (Rozbicki et al. 2015; Bhatta et al. 2017). Lu et al. (1997) indicated that protein content and wet gluten content of wheat increased in a paulownia/wheat intercropping system, compared to control. Wang et al. (2015) also reported that wheat starch and crude fat content were increased in apricot-based intercropping systems, and wheat grown nearer to apricot was poorer in quality. In this study, we found that tree shading significantly increased the protein content and wet gluten content (Table 4). These results are largely consistent with previous studies and confirm that tree shading during grain development can have a negative effect on grain yield but positive on grain quality.
In order to increase grain production, appropriate management measures are needed to minimize competition between trees and crops in intercropping systems. We suggest several specific recommendations to reduce competition in fruit tree-crop intercropping systems: (1) selecting crop and fruit tree varieties (e.g., shade-tolerant crop varieties, low-height fruit tree varieties, and rootstocks) that are more suitable for agroforestry systems; (2) appropriately increasing planting distances between and within the tree rows, and conducting regular pruning of fruit tree crowns to decrease shading intensity. This study only investigated the influence of apricot tree shade intensity on the grain yield and quality of intercropping wheat, but ignored the performance of the apricot trees. As a result, this study can be used for understanding limitations of wheat growth in apricot tree-based agroforestry systems, but it cannot be used to evaluate the overall performance of agroforestry systems.
Conclusion
Agroforestry is an effective approach for land use, which is widely adopted by farmers in Southern Xinjiang, Northwest China. The PAR reaching the crop canopy and TPAR (the PAR transmitted by fruit trees and wheat canopy getting to the ground surface), photosynthesis rate, and the total N and P contents of intercropping wheat were significantly decreased compared to monoculture wheat. The fruit tree shading affected the development of florets and spikelets, resulting in fewer grains per spike; the apricot tree shading had a negative effect on grain yield but positive on grain quality. Further research is needed to focus on shade-tolerant crop varieties and on how regular pruning of tree canopy may improve crop productivity in such systems.
References
AACC International (2010) Approved methods of analysis, 11th ed. AACC International, St Paul. http://methods.aaccnet.org/toc.aspx
Alam M, Olivier A, Paquette A, Dupras J, Revéret J, Messier C (2014) A general framework for the quantification and valuation of ecosystem services of tree-based intercropping systems. Agrofor Syst 88:679–691
Artru S, Garré S, Dupraz C, Hiel M, Blitz-Frayret C, Lassois L (2017) Impact of spatio-temporal shade dynamics on wheat growth and yield, perspectives for temperate agroforestry. Eur J Agron 82:60–70
Bergeron M, Lacombe S, Bradley RL, Whalen J, Cogliastro A, Jutras M, Arp P (2011) Reduced soil nutrient leaching following the establishment of tree-based intercropping systems in eastern Canada. Agrofor Syst 83:321–330
Bhatta M, Regassa T, Rose DJ, Baenziger PS, Eskridge KM, Santrad DK, Poudel R (2017) Genotype, environment, seeding rate, and top-dressed nitrogen effects on end-use quality of modern Nebraska winter wheat. J Sci Food Agric 97:5311–5318
Chirko CP, Gold MA, Nguyen PV, Jiang JP (1996) Influence of direction and distance from trees on wheat yield and photosynthetic photon flux density (Qp) in a Paulownia and wheat intercropping system. For Ecol Manag 83:171–180
Cui H, Jin L, Li B, Dong S, Liu P, Bin Zhao, Zhang J (2013) Effects of shading on dry matter accumulation and nutrient absorption of summer maize. Chin J Appl Ecol (Chin Engl Abstr) 24(11):3099–3105
Evers AK, Bambrick A, Lacombe S, Dougherty MC, Peichl M, Gordon AM, Thevathasan NV, Whalen J, Bradley RL (2010) Potential greenhouse gas mitigation through temperate tree-based intercropping systems. Open Agric J 4:49–57
Fixen PE, Grove JH (1990) Soil testing and plant analysis, 3rd edn. American Society of Agronomy and Soil Science Society of America, Madison
Friday JB, Fownes JH (2002) Competition for light between hedgerows and maize in an alley cropping system in Hawaii, USA. Agrofor Syst 55:125–137
Gao L, Xu H, Bi H, Xi W, Bao B, Wang X, Bi C, Chang Y (2013) Intercropping competition between apple trees and crops in agroforestry systems on the Loess Plateau of China. PLoS ONE 8:e70739
Gardner JS, Hess WM, Trione EJ (1985) Development of the young wheat spike: a SEM study of Chinese spring wheat. Am J Bot 72:548–559
Hong Y, Heerink N, Jin S, Berentsen P, Zhang L, Werf WVD (2017) Intercropping and agroforestry in China—current state and trends. Agric Ecosyst Environ 244:52–61
Inurreta-Aguirre HD, Lauri P-É, Dupraz C, Gosme M (2018) Yield components and phenology of durum wheat in a Mediterranean alley-cropping system. Agrofor Syst 92:961–974
Jaiswal CS, Mishra VK, Verma KS (1993) Intercropping ginger and turmeric with poplar (Populus deltoides ‘G-3’ Marsh). Agrofor Syst 22:111–117
Jose S, Gillespie AR, Seifert JR, Mengel DB, Pope PE (2000) Defining competition vectors in a temperate alley cropping system in the mid-western USA. 3. Competition for nitrogen and litter decomposition dynamics. Agrofor Syst 48:61–77
Kittur BH, Sudhakara K, Kumar BM, Kunhamu TK, Sureshkumar P (2016) Bamboo based agroforestry systems in Kerala, India: performance of turmeric (Curcuma longa L.) in the subcanopy of differentially spaced seven year-old bamboo stand. Agrofor Syst 90:237–250
Long SP, Zhu GX, Naidu SL, Ort DR (2006) Can improvement in photosynthesis increase crop yields? Plant Cell Environ 29:315–330
Lu Q, Yang H, Ci L, Zhu Z, Wu Y, Jing Y (1997) Effect of radiation transmission on crop yield and quality. Acta Ecol Sin (Chin Engl Abstr) 17(1):36–44
Makumba W, Akinnifesi FK, Janssen B, Oenema O (2007) Long-term impact of a gliricidia-maize intercropping system on carbon sequestration in southern Malawi. Agric Ecosyst Environ 118:237–243
Nasehzadeh M, Ellis RH (2017) Wheat seed weight and quality differ temporally in sensitivity to warm or cool conditions during seed development and maturation. Ann Bot 120:479–493
National Bureau of Statistics of China (2013) China Statistical yearbook. China Statistics Press, Beijing (in Chinese)
Peng XB, Zhang YY, Cai J, Jiang ZM, Zhang SX (2009) Photosynthesis, growth and yield of soybean and maize in a tree-based agroforestry intercropping system on the loess plateau. Agrofor Syst 76:569–577
Reynolds PE, Simpson JA, Thevathasan NV, Gordon AM (2007) Effects of tree competition on corn and soybean photosynthesis, growth, and yield in a temperate tree-based agroforestry intercropping system in southern Ontario. Can Ecol Eng 29:362–371
Rivest D, Lorente M, Olivier A, Messier C (2013) Soil biochemical properties and microbial resilience in agroforestry systems: effects on wheat growth under controlled drought and flooding conditions. Sci Total Environ 463:51–60
Rowe EC, Noordwijk MV, Suprayogo D, Hairiah K, Giller KE, Cadisch G (2001) Root distributions partially explain 15N uptake patterns in gliricidia and peltophorum hedgerow intercropping systems. Plant Soil 235:167–179
Rozbicki J, Ceglińska A, Gozdowski D, Jakubczak M, Cacak-Pietrzak G, Mądry W, Golba J, Piechociński M, Sobczyński G, Studnicki M, Drzazga T (2015) Influence of the cultivar, environment and management on the grain yield and bread-making quality in winter wheat. J Cereal Sci 61:126–132
Toyota M, Tsutsui I, Kusutani A, Asanuma K (2001) Initiation and development of spikelets and florets in wheat as influenced by shading and nitrogen supply at the spikelet phase. Plant Prod Sci 4:283–290
Wang J, Zhang Y, Liu C, Tan Y, Zhang Y, Chen L, Zhu H, Chen Y (2015) Wheat grain filling characteristics and quality traits in wheat-apricot intercropping field in Southern Xinjiang. Acta Agric Boreal Occident Sin (Chin Engl Abstr) 24(7):44–50
Willey RW, Holliday R (1971) Plant population, shading and thinning studies in wheat. J Agric Sci 77:453–461
Yang B, Gong P, Che Y, Zhang P, Xu Y, Yang L (2009) Study on almond and cotton intercropping to yield of cotton. Chin Agric Sci Bull (Chin Engl Abstr) 25(17):93–97
Yang L, Ding X, Liu X, Li P, Eneji AE (2016) Impacts of long-term jujube tree/winter wheat–summer maize intercropping on soil fertility and economic efficiency—a case study in the lower North China Plain. Eur J Agron 75:105–117
Zhang W, Wang BJ, Gan YW, Duan ZP, Hao XD, Xu WL, Lv X, Li LH (2017) Competitive interaction in a jujube tree/wheat agroforestry system in northwest China’s Xinjiang Province. Agrofor Syst 91:881–893
Zou X, Sanford RL (1990) Agroforestry systems in China: a survey and classification. Agrofor Syst 11:85–94
Acknowledgments
This work was funded by the National Natural Science Foundation of China (Grant Nos. 31560587, 31660370, 31760364), Special Fund for Agro-scientific Research in the Public Interest (201503116), and the Modern Agricultural Industry Technology System (CARS-03-49).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Qiao, X., Chen, X., Lei, J. et al. Apricot-based agroforestry system in Southern Xinjiang Province of China: influence on yield and quality of intercropping wheat. Agroforest Syst 94, 477–485 (2020). https://doi.org/10.1007/s10457-019-00412-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-019-00412-5