
Abstract
In recent decades, significant changes have been observed in the atmospheric pollutant emissions and deposition in the mountain regions of Central Europe. Pollution caused significant deforestation, but the level of tree damage differs, even between neighboring locations. Thus, it is particularly important to examine the relationships between pollutant deposition rate and detrimental changes in the subalpine spruce ecosystems, and to correlate these with the intensity and structure of pollutant deposition. Radial wood cores were extracted at breast height from trees located in different montane areas: the Sudetes and both Western and Eastern Carpathians. Specific features of the secondary xylem, i.e., annual ring width as well as the width and lumen of early- and latewood tracheids, were analyzed for three decades: before the intense of environmental pollution (1950–1960), during essential stage of the ecological disaster (1980–1990) and after cessation of pollution (2000–2010). The narrowest annual rings and earlywood tracheids were formed in the 1980s in areas where fog was the main source of the pollutant deposition. In conclusion, it has been shown that fog could be an important factor increasing the destructive role of pollutants on wood formation. It was manifested in decreasing of cambial activity resulting in formation of narrower annual rings as well as in reduction in radial dimensions of earlywood tracheids.
Similar content being viewed by others
Introduction
The Sudetes, a mountain range in southwestern Poland, are characterized by Norway spruce (Picea abies) forest ecosystems. Approximately 30 years ago, acid deposition caused an unprecedented environmental disaster in the Western Sudetes (Abrahamson et al. 1989; Acker et al. 1995; Matschullat et al. 2000). This was exacerbated by the Norway spruce monoculture and massive deforestation that occurred in this period, increasing the fragility and low hazard resistance of the forest. In this context, high air pollution levels have become the main factor in weakening resistance (National Institute for Public Health and Environment 2010; Nilsson and Grennfelt 1988; Schweingruber 1996).
Poland, Czech Republic and Germany were in the group of European countries with the largest sulfur and nitrogen emissions (Vestreng et al. 2007). Up until 1970, the long-term upward tendency in the emission of SO2 in the region of “the Black Triangle” (the area of the Polish–Czech–German borderland) was more or less similar to that observed throughout the whole Central-Eastern Europe. Between 1970 and 1990, however, a sharp increase in pollutant deposition was observed and reached an unprecedented value of over 800 mMol m−2 year−1 at its peak (Kopáček and Veselý 2005). In 1950, the burning of lignite coal for electric energy had already 40% share, and jointly with coal 97%. The emission was dominated by the collection of high stacks, so it was subjected to the long-range transport. Much smaller emission of this kind was observed in Ukraine (Szoege 2007). Before the year 2000, emission of air pollutants has decreased substantially as a result of commitments imposed on the countries by protocols to the convention on long-range transport of air pollutants (CLRTAP). Over the last decades, abatement in SO2 emissions reached 80% over the period 1980–2016 and 48% if NOx is considered (EMEP 2018).
Pollution can be characterized by dry, wet, or fog deposition, depending on the pathway of pollutant flux to the environment. Direct deposition of fog to vegetation, called also “horizontal precipitation,” means that tiny fog droplets meet any receptor present. This is particularly efficient when fog is accompanied by strong wind. Such conditions are frequent in low clouds with bases below the level of existing mountain summits and ridges. Pollutant concentrations in clouds or fog are usually several times higher than in atmospheric precipitation (Błaś et al. 2012; Pahl et al. 1994; Weathers et al. 1995). For example, at elevations higher than 800 m a.s.l. in the Black Triangle, fog moisture input causes atmospheric pollutant deposition to increase (Acker et al. 1995; Błaś et al. 2010; Matschullat et al. 2000), highlighting its importance as a pathway for forest destruction, especially in the medium-height central European mountain ranges (Sudety Mts., Ore Mts., Bohemian Forest, Harz etc.). Landuse type (e.g., grass, forest, rocks) influences pollutant deposition via fog causing huge local contrast of fog precipitation and pollutant deposition (Błaś et al. 2010; Lovett 1984; Sobik 1991; Weathers et al. 1995). Taking height and the surface area index (SAI) into consideration, the highest horizontal precipitation efficiency is observed for spruce trees (Klemm and Wrzesinsky 2007). Typically, within individual mountain ranges in Poland, precipitation totals show limited differences between 80 and 120% of the aerial average. On the other hand, fog water deposition in the same area may differ, respectively, from 10% to even 1000% (Archive 2018; Błaś et al. 2008). The largest volume of fog water is deposited at high elevated sites exposed to winds from the western sector which cause the formation of dense orographic fog. Therefore, close to upper treeline in the Sudetes, total pollutant deposition was composed of around 30–40% by precipitation, 20–25% by dry deposition and 35–50% by direct fog/cloud deposition, while the associated proportion at the nearby lowland sites were around 75%, 20% and 5%, respectively (Błaś et al. 2008). The spatial diversity of fog deposition and pace at which montane forest ecosystems are destroyed have been examined by comparing daily pollutant loads according to the atmospheric circulation direction (Godek et al. 2012). Accordingly, it has been shown that on a mountain top in the Sudetes, the wind from the SW–NW sector contributes as much as 66% of deposition, with the wind from the N–NE sector forming a secondary maximum (23% of deposition), while only 5% is deposited during airflow from the wide E–SE–S range (Godek et al. 2012). This indicates a high polarization in pollutant deposition via fog, depending on slope exposure to a given directional sector.
Air pollutants negatively influence forest tree growth, which is manifested in changes to the crown shape (de Kort et al. 1991), tree defoliation (Kurczyńska et al. 1997), and decreasing resistance to drought, frost, and pathogens (Caporn 2013). In polluted areas, metabolic variations are caused by transpiration and photosynthesis disorders (Bytnerowicz 1996; Jadczyk 2009; L’Hirondelle and Addison 1985). Ultimately, even in mildly polluted areas, forest decline is an evidence of increasing tree mortality (Linzon 1971). In the 1980s, a massive spruce forest dieback began, affecting approximately 42% of coniferous forests in the Sudetes (Zawiła-Niedźwiedzki 1994).
Since the 1970s, scientific research has started to focus on the impact of air pollution on wood formation, mainly treated as a modification of annual rings width, which is related to the divisional activity of cambium and the further expansion of newly formed cells (e.g., Dünisch et al. 1996; Evertsen et al. 1986; Kurczyńska et al. 1997; Linzon 1971; Nash et al. 1975). Cambium is a cylindrical lateral meristem, which is located beneath the bark of trees and shrubs and gives rise to the secondary conductive tissues: xylem (wood) and phloem (Evert 2006). The processes of cambial activity and secondary tissues formation are controlled by intrinsic factors, e.g., tree age, hormones, and gene expression (Aloni 2015; Hertzberg et al. 2001; Schrader et al. 2004; Uggla et al. 1996; Zajączkowski and Wodzicki 1978). However, trees are also exposed to environmental factors, both climatic (e.g., temperature and drought) (Begum et al. 2018; Castagneri et al. 2017; Sidor et al. 2015; Wodzicki 2001) and non-climatic (e.g., forest density, pests, and extreme events) (Bräuning et al. 2016; Ferretti et al. 2002; Juknys et al. 2014).
The cambium as a meristematic tissue functions over the years and is influenced by the environmental factors. The cambial cell divisions, both producing the derivative secondary tissue and increasing the cambium circumference, as well as formation of new rays, etc., are consistently recorded upon cambium activity in the wood structure. Therefore, the history of all the cambial events can be traced in successive annual rings (Bannan 1950; Carlquist 1988; Hejnowicz 1971; Myśkow and Zagórska-Marek 2004, 2013). This is a common method used in wood anatomy and dendrochronology for the analysis of, for instance, the impact of fluctuations in environmental factors on the wood structure.
Numerous analyses have revealed a sharp decrease in annual ring width in Picea abies growing in the following polluted areas: high-altitude stands of the Erzgebirge (Dünisch et al. 1996), in the Sudetes (Godek et al. 2012; Rydval and Wilson 2012; Kolář et al. 2015), Ore Mts. (Wimmer 2002; Samusevich et al. 2017), Lithuania (Juknys et al. 2014) and southern Germany (Elling et al. 2009). The scale of this reduction provides information on pollutant persistence and impact.
Forest ecosystem degradation is characterized by the coexistence of completely or moderately deforested areas alongside neighboring forest stands remaining in a relatively good condition; the spatial diversity of wet deposition via atmospheric precipitation is insufficient in explaining the reasons for this. It is more important to understand both the spatial diversity of fog deposition, and the period for which trees are exposed to fog in the form of heavily polluted low clouds (L’Hirondelle and Addison 1985).
The aim of the present work was therefore to determine differences in tree growth impairment related to spatial and temporal differences in pollutant deposition at the studied areas, and to answer the following sequence of questions:
-
Does the high level of pollutant deposition result in decreases in tree-ring width, and in other changes in the secondary xylem?
-
Does tree-ring width in Norway spruce show a clear response to spatial changes in atmospheric pollutant deposition?
-
Is the spatial pattern of tree-ring reduction related to fog deposition more than to other deposition components?
Although the acid deposition-induced forest decline tends to be a past problem in Europe and North America, the profound understanding of the role of primary factors controlling the deposition could help to take measures against ongoing forest degradation processes in other countries, e.g., in Eastern Asia (Igawa et al. 2015; Shigihara et al. 2008; Shimadera et al. 2011; Song et al. 2017; Yamaguchi et al. 2015).
Materials and methods
Selection of locations and climate conditions
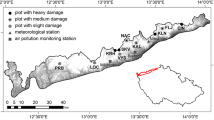
Research was performed on Norway spruce (Picea abies) growing in three mountain ranges: the Sudetes, the Western Carpathians and the Eastern Carpathians (Fig. 1). The sequence of 12 sampling sites was designed along the NW–SE profile line, parallel to the prevailing direction of atmospheric circulation of maritime origin. The selected trees were growing on similar climatic and orographic conditions in terms of slope inclination. Soil cover in all sites was highly skeletal throughout the profile and dominated by Dystric Cambisols and Eutric Cambisols. In the Carpathians, climate is slightly more continental than in the Sudetes, which is expressed by 1 °C larger annual temperature range and similar annual precipitation (Table 1). Exposure to advection of humid Atlantic air masses and the absence of other mountains on a windward side are the reasons why the higher frequency of fog and horizontal precipitation sums are observed in the Sudetes (Table 1).

Geographical setting of the study area and location of sampling sites (see also Table 2)
The sampling sites were selected as references for different levels of air pollution. The criteria adopted included an appropriate distance from emission sources as well as the role of pollutant fog deposition (Table 1). In case of high emission sources, pollutant deposition pattern is not a simple function of a distance from the source due to the efficiency of atmospheric mixing, transport and deposition mechanisms (see comparison between two locations at the Szrenica Mt.—Table 1). Meanwhile, tens or even hundreds of kilometers further, on a windward orographic barrier the deposition rate may increase rapidly as a result of pollutant washout (Lysá Hora—Table 1). A cause of this may be higher rainfall and significant share of fog pollutant deposition (Błaś et al. 2002; Dore et al. 1992, 1999; Pahl et al. 1994; Weathers et al. 1995). To indicate precisely the role of pollutant deposition via fog, several measurement campaign were performed and data published (Błaś et al. 2012; Godek et al. 2012; Kryza et al. 2009; Skybova 2006). Significantly larger total pollutant deposition below forest canopy (throughfall) as well as fog contribution in total deposition was observed at the westernmost Szrenica Mt. (Sudety Mts.) than more to the southeast at Lysá Hora (Wester Carpathian) and Płaj (Eastern Carpathian; Table 1). These differences might be explained by more intense orographic deformation of predominant airflow from the western sector, higher frequency and liquid water content of fog and finally strong influence of the atmospheric sources of pollutant emission in the vicinity of the Szrenica Mt. (Archive 2018; Błaś et al. 2012; Černy 1994; Kulhavy and Klimo 1998; Oulehle et al. 2010; Skybova 2006).
Therefore, forest ecosystem degradation is characterized by the coexistence, in close vicinity to each other and exhibiting similar site conditions, of totally deforested areas, areas of moderate deforestation, and forests in a relatively good condition (Zawiła-Niedźwedzki 1994). For that very reason, authors decided to vary the sampling location in each study group (A, B and C). They are not necessarily organized in accordance with geographical distribution:
Group A Outside the area of exceedance of acid critical loads in Europe, very low to low fog pollutant deposition (Table 2); windward site in the upper subalpine forest zone; medium-dense mixed forest with Norway spruce; trees have well-developed crowns without visible injuries; reference point.
Group B Low to medium of exceedance of acid critical loads as well as fog pollutant deposition (Table 2), valley or south-eastern slopes; lower subalpine forest zone ecosystem; dense Norway spruce forest; tree injuries are rare; dead tree trunks below 10%.
Group C High and very high exceedance of acid critical loads as well as fog pollutant deposition (Table 2), ridges or western and northwestern slopes; upper subalpine forest zone ecosystem, extreme thinning of Norway spruce forest; comprises 90% dead tree trunks, and only individual living trees.
Dendrochronological analyses
All 286 core samples used in the present study were collected between May and October 2013 from 12 locations and divided into the three groups, as described above (Fig. 1). At each location, 16–29 core samples were collected (Table 2) at the breast height, using an increment borer (MORA CORETAX, Haglöf, Sweden), with the auger set to 400 mm in length. This is in accordance with studies of tree population dynamics or short-term ecological assessments (e.g., stand growth declines) which usually involve extracting one core per tree as well as 20–30 trees in a single location (Fritts 1976; Norton 1998; Schweingruber 1988). Trees that were the most representative of the site were selected for sampling, according to the following criteria: old enough at least (70–80 years), trunk circumference, tree height, crown habit, degree of injury, stand density, and substratum type around the tree. The sampling intensity varied between sites due to limited numbers of living trees on ridges (heavy deforestation), but also small contribution of trees at the right age and species.
After high-resolution scanning (LA2400 scanner; Fig. 2), dendrochronological samples were subjected to analysis using the semi-automatic image analysis program WinDENDRO V.2016a (Regent Instruments Canada Inc 2009). Like other image analysis programs, WinDENDRO detects annual and intra-annual ring boundaries using changes in image light intensity, but manual adjustment to the ring boundaries is required to account for growth anomalies. To assure the proper quality and to eliminate the age factor in the gathered data, the measured tree-ring width values were transformed through quality checking with COFECHA software (Grissino-Mayer et al. 1996) as well as the ARSTAN program (Cook and Holmes 1986), in which the measurement series were individually detrended using a cubic smoothing spline with a 50% frequency response cutoff equal to 67% of the series length. Next, the ratios between ring widths and the fitted curves were calculated as detrended series. A multiple regression model was used to describe the climate relations with tree growth, but recent evidence suggests a more complex and nonlinear relationship between tree growth and climate (Wilmking et al. 2005; Lloyd et al. 2013; Lee et al. 2016). Elling et al. (2009) documented that temperature limited trees can become insensitive to temperature as a result of pollution load. Decreased temperature response may be a consequence of pollution-related limitation of photosynthesis through nutrient deficiency or direct foliage damage.

Outline of analyzed cores: a reference site, core no. 2; b site with limited fog deposition, core no. 4; and c site with intense fog deposition, core no. 11. Analyzed annual rings are denoted by colored rectangles: 1950–1960 blue; 1980–1990 red and 2000–2010 green (color figure on line)
Anatomical analyses
After completing the dendrochronological analyses, the second part of the research was conducted on the basis of the obtained results, aimed at the detailed anatomical studies of wood formed in trees growing in the analyzed 12 locations. Due to high internal correlation of individual cores within any location, for each of them one core with the age of 70–80 years and the strongest correlation with the master series was selected. Thus, such a core was as representative and similar to trees growing in the same location as possible.
Figure 1 and Table 2 show the tree sampling sites and the tree characteristic features. Since the level of air pollution was changed during the 20th century, to analyze the impact of air pollution on wood formation we had to compare the selected ring formed in specific years with the aim to avoid the ring with transient character. Thus, three periods (decades) recorded in the annual rings were selected per core for the anatomical analysis:
-
1950–1960: rings formed before the increase in environmental pollution; reference rings.
-
1980–1990: rings formed during the period of intense air pollution.
-
2000–2010: rings formed after cessation of pollution; comparative period.
The cores were divided into fragments 20–50 mm long, with selected annual increments. Transverse sections (30 μm thick) were cut from the fragments on a sliding microtome Leica S14 2004 (Leica Instruments, GmbH, Germany; Fig. 3). The sections were processed according to standard protocols for wood material (Myśkow 2010; Myśkow and Zagórska-Marek 2013; O’Brien and McCully 1981).

Representative transverse sections of selected annual rings. The core no. 11 illustrates widths in relation to intense fog pollutant deposition. All sections are presented at the same scale (bar 500 µm). a 1950–1960 (reference period), one ring formed before the increase in environmental pollution; b 1980–1990, several rings formed during the period of intense pollutant deposition, and conspicuous severe decrease in annual increments; c 2000–2010 (comparative period), the annual ring width increased after cessation of pollutant deposition
Annual ring width, and the width and lumen of selected tracheids were measured for each of the three periods (1950–1960; 1980–1990 and 2000–2010). Three radial rows of tracheids were randomly chosen from each annual ring, and 14 cells were measured from each row (42 tracheids per ring), comprising the first tracheid formed at the beginning of the growing season, six earlywood cells, and seven latewood cells, formed at the end of the growing season. The only tracheids located at the opposite margins of the annual rings were selected to indicate the value of features representative for early- and latewood tracheids. This method was repeated for all annual rings in each decade, so over 400 cells were measured per decade. The large number of cells as well as obtained appropriate standard error indicated that it was sufficient for further statistical analyses. In very narrow rings, all tracheids were measured: they were classified as early- or latewood tracheids, according to the criteria of Mork (1928) and Denne (1988), by which a latewood cell is defined as a cell in which total wall thickness multiplied by two is larger than or equal to the lumen width. Cell wall thickness was calculated from the measured cell widths and lumens. For all measurements, average values and standard errors were then calculated for all cell properties in each decade. Data were analyzed and documented using an Olympus BX50 Microscope and Olympus Camera, DP71 using CellˆB Software (Olympus Optical). Measurements were performed using AxioVision LE (Carl Zeiss Vision GmBH), and values were calculated in Excel 2016. CorelDRAW 2017 (Corel Corporation) was used for processing the digital images.
Statistical analyses
Statistica version 13 software (StatSoft, Inc., 2014) was used for data analyses. Data normality was assessed using Shapiro–Wilk’s W-test. In cases of non-normal distribution, the data were transformed using log transformations. Levene’s test was then used to test homogeneity of variances. The differences between cell width and lumen between the three periods (decades) in the three areas were compared using a one-way parametric ANOVA. Welch’s t test was used to correct any unequal variances between groups. When an ANOVA revealed significant effects at P < 0.001, a post hoc Tukey’s test was used to indicate the differences between analyzed groups (Zar 1998).
Results
In the present study, 12 measurement locations, representing three area groups (Groups A, B, and C; Table 2) were selected.
Group A: limited wet and fog pollutant deposition
Dendrochronological analysis
Group A (the reference group) consisted of two sites (Fig. 1, Table 2), from which two groups of trees were chosen for analyses (Groups 1 and 2). These locations had different elevations and climatic conditions, being a lowland (262 m a.s.l.) and mountain site (1340 m a.s.l.), respectively. However, the point-reunification criterion was the rate of fog and wet pollutant deposition, which is not as intense across the Eastern Carpathians in comparison with the Sudetes.
The site for Group 1 (21 trees) was located in the foothills of the Eastern Carpathians, and Group 2 (25 trees) was situated much higher little below the tree line. The full sequence of annual increments indicated that the mean growth rate was 1.57 and 2.04 mm for Groups 1 and 2, respectively (Table 2). A lapsing trend for tree-ring width was expected in 1948–2012, due to increases in tree diameter (Fig. 4a). However, increasing trends were also observed in 1966–1970, 1994–1997, and 2006–2011. After removing the age-related growth trend (Fig. 4b), the tree-ring width standard chronology (STD) was relatively stable before the 1980s, with weak interdecadal variability. The variability in STD was enhanced after the 1980s, with a 20% decline in growth from the 1980s to 1990s, and a peak at the end of the series (20% above STD equilibrium).

Time series of a raw tree-ring width chronology; b tree-ring standard chronology (STD) in Norway spruce, in the form of a 5-year moving average of annual increments for the three analyzed groups; c reconstructed trends in S–SO42− deposition in area of the Polish–Czech–German borderland (from Kopáček and Veselý 2005) and annual average air temperature at the Szrenica Mt. (the Sudetes, Archive 2018)
Anatomical analysis
Measurements were made of the intrinsic wood properties recorded in the annual rings for the three selected decades (1950–1960; 1980–1990; 2000–2010). Two cores (nos. 1 and 2) representing the first and second groups of cores were selected for detailed anatomical analysis. The annual ring widths for each decade were comparable between cores, which was clearly visible on the surface of the cores (Fig. 2a, Table 3). A slight decrease in incremental width related to tree age was observed, especially in 1980–1990, similar to other Group A cores (Fig. 4b, Table 2).
The analysis of cell features in both cores showed that earlywood and latewood cell widths were homogenous (Fig. 5a, Table 3). The ANOVA showed a slight difference (F = 7.58, P < 0.0001) in the earlywood tracheid cell lumen between 1980–1990 and 2000–2010. Similarly, the latewood tracheid cell lumen formed in 1950–1960 showed differences (F = 11.639, P < 0.0001).

The width and lumen of early- and latewood tracheids analyzed in three decades for selected groups of trees (Groups A, B and C). Significant differences in mean values (shown in µm) between studied decades were tested by a one-way parametric ANOVA and are denoted by lower case letters. Whisker plots mark the standard errors
Group B: intense wet pollutant and limited fog pollutant deposition
Dendrochronological analysis
Group B comprised seven locations situated in the lower subalpine zone and leeward slope (eastern exposure) of the Sudetes and Western Carpathians (Fig. 1), from each of which 16–29 cores were collected; 170 tree cores were analyzed in total (Table 2). The full sequence of annual increments indicated changes in the mean growth rate at these locations in the range of 1.32 and 3.88 mm (Fig. 4a). From the beginning of the 1970s, there was a marked trend of decreasing annual ring size, which persisted until the mid-1980s, a period which overlapped with the gradual increase in pollutant emission and deposition. Thus, the 1950s have been assumed as the reference point for further consideration of trees vitality. At the peak of the ecological disaster (1980–1990), a decrease in annual increments of around 35%, compared with the 1950–1960 values, was recorded (Fig. 4b). The resulting slightly increased defoliation, in comparison with Group A, led to a decline in growth, and sometimes missing rings (Table 2). After the 1980s, a systematic improvement in tree vitality was observed; in 2000, the annual increment had approximately reached values recorded in the 1950s.
Anatomical analysis
Seven Group B cores (nos. 3–9) were selected for analysis, one from each location. Some specimen-specific modifications in ring width and wood cell parameters were observed (Table 3). Additionally, core no. 9 was collected from a mechanically destroyed tree, in which there was visible damage to the upper part. A decrease in annual ring widths was observed in 1980–1990 in all anatomically analyzed cores and was most pronounced in core no. 3. In 2000–2010, the annual ring width increased in comparison with 1980–1990, a finding that accorded with the STD of Group B cores (Fig. 4b; Tables 2 and 3). The reduction followed by increase of annual ring widths was also clearly visible macroscopically on the surface of wood cores (Fig. 2b). An exception was noted for cores nos. 3 and 5, in which a tendency of only a slight or no decrease in annual ring width was observed. In addition, in core no. 9, from the damaged tree, the annual ring width distinctly decreased in 2000–2010 (Table 3).
Measurements made of the early- and latewood tracheids in core nos. 3–9 are shown in Fig. 5b and Table 3. The ANOVA highlighted a significant difference between the width (F = 54.104, P < 0.0001) and lumen (F = 46.656, P < 0.0001) of earlywood tracheids formed during 2000–2010 and those formed during the other two periods (1950–1960 and 1980–1990). A slight difference was also visible in the latewood tracheid cell width. In these cases, values were lowest (F = 26.295, P < 0.0001) in 1950–1960. Data relating to the latewood cell lumen were homogenous between decades. Core no. 9 was excluded from the 2000–2010 analysis, because of the visible tree damage and changes in the wood cell parameters; however, average values were calculated using all (seven) analyzed cores for the decades 1950–1960 and 1980–1990.
Despite the comparable results for the Group B cores, they also presented a specimen-dependent modification of the parameters (Table 3). For example, in 1980–1990, core no. 3 was characterized by the lowest annual growth increments, and narrowest earlywood tracheids and lumens. Furthermore, cores no. 7 and 8 showed no increases in width and lumen in 2000–2010, in contrast to cores no. 3–6. In addition, annual rings from core no. 9 for 2000–2010 differed in terms of a significant reduction by 12–13 µm in earlywood tracheid widths and lumens (Table 3).
Group C: Intense wet and fog pollutant deposition
Dendrochronological analysis
Group C comprised three locations (nos. 10–12), in the westernmost part of the Sudetes and the Western Carpathians (Moravian–Silesian Beskids; Fig. 1, Table 2). A total of 28, 24, and 18 cores were collected for dendrochronological analysis from location nos. 10, 11, and 12, respectively (Table 2). The absolute values in mm showed that, over the whole study period, the narrowest annual rings were in Group C with a mean growth rate between 1.04 and 1.79 mm (Fig. 4a, Table 2). This result was expected due to the higher altitude compared with Groups A and B, characterized by a shorter vegetation period, longer snow cover period, higher wind speed, lower sunshine duration, and ice intensity. In addition, a stable decreasing tendency and largest reduction in tree vitality was observed at all Group C sites, which were well exposed to the prevailing wind direction in the westernmost parts of the Sudetes and Western Carpathians.
Dendrochronological investigations determined there to be an intense decrease (67%) in tree-ring width between the 1950s and 1980s (Fig. 4b); the most intensive phase of forest destruction was in 1980–1990. In the most extreme case (location no. 11), the mean tree-ring width decreased by as much as 80%, corresponding to an extreme worsening in ecosystem health that approached complete degradation. Minimum tree-ring widths during the most intensive phase of the ecological disaster were as low as 0.1–0.2 mm. Therefore, instances of missing rings were more numerous in Group C than in Groups A and B. Sporadic occurrences of missing rings were found in the earlier chronology, but a dramatic increase was observed during 1978–1986, when 89% of such cases took place. The number of missing rings started to reduce after 1992, and none of the analyzed trees had missing rings in the last 20 years (Table 2). Similar results have been observed by Kolář et al. (2015), Samusevich et al. (2017) and Treml et al. (2012).
Cores were collected in 2013 and were taken exclusively from the trunks of live spruce, which had survived the ecological crisis that ended 20 years earlier. From the beginning of the 1990s, the results for Group C correlate with the observed systematic improvement in forest health, mainly related to sites where the earlier vitality decrease had been most marked. In 2000–2010, tree-ring width at these sites reached 140% of the value recorded for 1950–1960 (Fig. 4, Table 2).
Anatomical analysis
Three representative cores (nos. 10–12) were selected from Group C (Tables 2 and 3). The most pronounced decrease in annual ring width was noted in 1980–1990, compared with the other two decades, with wood ring width differing from 70 to 735 µm, and sometimes only 3–4 cells being formed during the vegetative season (Figs. 2c, 3 and 5c; Table 3). These data agreed with the tendency of decreasing STD in Group C (Fig. 4b). Additionally, traumatic resin ducts were observed in the narrow annual rings.
Figure 5c and Table 3 show the results of the anatomical analyses and early- and latewood cell measurements. The ANOVA determined large, significant differences between earlywood tracheid cell width (F = 256.654, P < 0.0001) and cell lumen (F = 256.598, P < 0.0001) in 1980–1990, compared with tracheids formed during the other two periods. This showed that, in the period of most intense air pollution, cell size (width and lumens) decreased by approximately 10 µm (24–29%), on average. The latewood tracheid parameters were homogenous in this group of cores (Fig. 5c, Table 3).
Discussion
Hauhs and Wright (1988) point out to the compounds of sulfur and nitrogen as the main agents responsible for acidification of the atmospheric precipitation. It needs to be stressed, however, that of the two, the single SO2 molecule is over twice as responsible for acidification of the environment (Bingman et al. 1989). In the area of the Polish–Czech–German borderland, the effects of environmental acidification and deforestation as a consequence reached the scale of the ecological calamity (Elling et al. 2009; Vacek et al. 2013). However, this disaster in the European highlands ended some 25 years ago and the extent of the damage has decreased considerably. Therefore, at present it is possible to observe not only the changes related to the forest damage, but also how the forests return to the vitality. Thus, the papers continue to be published, attempting to explain both why it occurred and its spatial context (Kolář et al. 2015; Samusevich et al. 2017; Treml et al. 2012).
It is generally accepted that exposure to pollutants negatively affects tree functioning in different ways (Caporn 2013; Emberson 2003). In order to determine the impact of air pollution on cambium activity, samples of Picea abies wood were analyzed and compared between three groups of locations, differing in levels of air pollution intensity. This therefore enabled changes over time in wood structure to be compared.
In highly polluted areas with a prevalent presence of fog/cloud, our results clearly showed a significant reduction in annual ring width (> 60%), with the concomitant formation of the narrowest earlywood tracheids, reduced by one quarter of their diameter in comparison with the other two decades (1950–1960 and 2000–2010). A reduction in annual ring width is a known response to high pollution levels, especially when trees grow near the pollutant source as was observed in Germany, the Czech Republic, Lithuania and Poland. In these places, the main toxic substances were: sulfur dioxide and nitrogen oxides (Dünisch et al. 1996; Juknys et al. 2014; Sensuła et al. 2015). Our results (Fig. 4) indicated the inverse proportionality between tree-ring standard chronology with reconstructed trends in wet-only S–SO42− deposition in area of the Polish–Czech–German borderland (Kopáček and Veselý 2005; Fig. 4c). While the peak of the S–SO42− deposition occurred in 1981, the deepest tree-ring width reduction started 2 years later in Groups B and C. In case of reference Group A which is placed in a large distance from pollutant emission sources, the reductions were not observed even in high elevated slopes exposed to the west and northwest. This means that reducing of the increment reaction of trees marked once 30 mMol m−2 yr−1 deposition S–SO42− has been exceeded (Fig. 4c).
Additionally, we showed that the largest wood parameter reductions in 1980–1990 occurred at sites with the highest frequency of severely acidified fog, connected with a low-lying cloud base of orographic origin (Błaś et al. 2012; Godek et al. 2012; Igawa et al. 2015; Shimadera et al. 2011; Yamaguchi et al. 2015). This concerned sites at the westernmost and northwesternmost edges of the Sudetes and Moravian–Silesian Beskids, which were also well exposed to the advection of air from the W–NW sector (Group C). The location of these sites on the NW edge of the Sudetes, i.e., the side exposed to the prevailing circulation and largest pollutant sources, means they are characterized by the highest fog efficiency and pollutant deposition (Błaś et al. 2002, 2012; Dore et al. 1999; Godek et al. 2012). The understanding of the reasons and the course of changes is facilitated by information on spatial diversity of the exposure time of the trees to heavily polluted atmospheric water. The cumulative time during which trees are exposed to heavily polluted fog water is around threefold longer (35–45%) than for precipitation (10–15%; Archive 2018).
At locations where the cloud base rarely covers the subalpine Norway spruce forest, the reduction in annual tree increments was limited (< 30–35%). Representatives of this group (Group B) were located in the SE–E of the Sudetes and Western Carpathians, as well as on slopes of SE–E exposure, i.e., the leeward slope in relation to the prevailing wind direction. The lower tree-ring reduction here was related to the gradual depletion and partial removal of pollutants by an air mass infiltrating the mountain region along a WNW–ESE profile. This results in a systematic reduction in wet deposition efficiency, and in reduced fog deposition; the latter is caused by a gradual rise in the orographic cloud base, as well as a reduction in both cloud liquid water content and the cloud pollutant load it transports. This is also confirmed by the lack of deforestation observed in the Eastern Carpathians (Group A).
Taking into account the anatomical analyses, despite the severe reduction in annual ring widths, we found that modifications to wood formed in highly polluted areas with fog mostly concerned earlywood tracheids, as previously noted by Dünisch et al. (1996) for the high concentration of sulfur dioxide in East Germany. On contrary, other authors observe modifications to latewood tracheids, resulting in decreases in wood density affected also mainly by sulfur dioxide in Ireland and Germany (Evertsen et al. 1986; Wimmer 2002).
The severe destruction of trees located in highly polluted areas subjected to intense fog pollutant deposition was also manifested in missing rings and the formation of traumatic resin ducts. These features in growth ring sequences indicate severe, stressful events (Dünisch et al. 1996; Wimmer 2002).
The final width of annual rings is an outcome of two interrelated processes. The first of these is cambium divisional activity: the number of wood cells formed during the vegetative season directly relates to the number of periclinal divisions of cambial cells. The second process relies on cell size, which is related to the growth and differentiation of wood cells produced in a particular annual ring (Aloni 2015; Wodzicki 2001). Both these processes were shown to be impaired during formation of the narrowest wood rings. Decreasing numbers of cambial cell divisions resulted in a reduced number of tracheids. Simultaneously, the narrowest cells originated because wood cell growth was limited in the radial direction, as noted by de Kort et al. (1991).
The reduction in pollution emissions in the mid-1990s translated into a systematic improvement in forest conditions at all sites, frequently reaching the level recorded in the reference period; this indicated the ability of trees to return to previous vitality levels, as has been observed in different tree species growing, e.g., in Poland, Lithuania, and the Czech Republic (Danek 2007; Juknys et al. 2014; Rydval and Wilson 2012). Both the earlier decline and present increase in health are significantly spatially diverse. The processes by which annual ring width increases and decreases respond sufficiently quickly to environmental stimuli to act as an important indicator of forest ecosystem degradation. In our study, Norway spruce showed a tendency for broadening of annual rings and wood cell parameters in 2000–2010, corresponding to improving environmental conditions. This was observed not only in trees located in areas with significant pollutant fog deposition, but also in Group B trees, which were characterized by high deposition levels but with only secondary participation of fog/cloud. In this case, after only moderate decrease of wood parameters in 1980–1990, there was a significant increase in the earlywood cell parameters in 2000–2010, possibly resulting from the cessation of intensive pollution. In addition, climate warming may induce secondary growth, especially in trees growing at high altitudes (Sidor et al. 2015). Both these factors may be responsible for the intensification of secondary growth observed in our samples.
The response of trees to negative or positive environmental factors is evident in the quantity and quality of wood formed in the annual rings (Bräuning et al. 2016; Wodzicki 2001). Additionally, due to differences in factors such as genotype and/or environmental (climatic, soil) microscale conditions, responses may be individual and specimen-specific (Juknys et al. 2014), as confirmed by our results. We are aware that ring width is affected not only by air pollution but also by environmental factors, such as drought, frost, precipitation, etc. (Ferretti et al. 2002; Nash et al. 1975). However, by comparing the anatomical data with the results of a broader dendrochronological analysis of over 200 cores, we were able to exclude these other factors from the analysis of significant reductions in ring width growth. Therefore, we could demonstrate that fog/cloud pollutant deposition significantly affects cambium activity and tracheid differentiation.
The consequences of air pollution for forests in the mountainous regions of Central Europe clearly seem to be limited in time and space, because of the spatial pattern of pollutant deposition with dominant role of fog deposition. The extent of the damage has decreased considerably during the last 25–30 years, and thus the improvement in forest vitality is observed nowadays.
Conclusions
The reduction in cambial activity manifested by narrow annual tree rings is a known feature of highly polluted areas. Our research confirmed the negative impact of air pollutants on cambium activity and secondary wood formation in Norway spruce. Moreover, during the period of highest pollutant emissions, distinct reductions in these parameters were observed at sites with the highest frequency of severely acidified fog. Simultaneously, in highly polluted locations but with limited fog occurrence, where trees more often remained below the cloud base, there was a less clear reduction in annual wood increments. The findings therefore indicated the role of the pollutant deposition pathway, i.e., wet vs. fog/cloud deposition, on the final pollutant impact. Deposition via particularly polluted fog/cloud droplets negatively influences both cambium activity and tracheid differentiation. Consequently, the reduction in the number and thickness of tracheids formed during the vegetative season results in the formation of narrow annual rings.
References
Abrahamson G, Seip HM, Semb A (1989) Long-term acidic precipitation studies in Norway. In: Lindberg SE, Page AL, Norton SA (eds) Acidic precipitation. Springer, New York, pp 137–145
Acker K, Moller D, Wieprecht W, Naumann S (1995) Mt. Brocken, a site for a cloud chemistry measurement programme in central Europe. Water Air Soil Pollut 85:1979–1984. https://doi.org/10.1007/BF01186124
Aloni R (2015) Ecophysiological implications of vascular differentiation and plant evolution. Trees 29:1–16. https://doi.org/10.1007/s00468-014-1070-6
Archive (2018) Meteorological observatory of the Department of Climatology and Atmosphere Protection. Wrocław University, Wrocław
Bannan MW (1950) The frequency of anticlinal divisions in fusiform cambial cells of Chamaecyparis. Am J Bot 37:511–517. https://doi.org/10.2307/2438026
Begum S, Kudo K, Rahman MdH, Nakaba S, Yamagishi Y, Nabeshima E, Nugroho WD, Oribe Y, Kitin P, Jin H-O, Funada R (2018) Climate change and the regulation of wood formation in trees by temperature. Trees 32:3–15. https://doi.org/10.1007/s00468-017-1587-6
Bingman I, Thunberg B, Berlekom M, Pleijel H, Agren Ch (1989) Ponadgraniczne Zagrożenie Środowiska. Fakty o zanieczyszczeniu powietrza oraz o zakwaszeniu. Szwedzko-Polskie Towarzystwo Ochrony Środowiska i Urząd Ochrony Przyrody, Sztokholm, p 44. (in Polish). http://dlibra.kul.pl/dlibra/doccontent?id=453
Błaś M, Sobik M, Quiel F, Netzel P (2002) Temporal and spatial variations of fog in the Western Sudety Mts. Poland. Atmos Res 64:19–28. https://doi.org/10.1016/S0169-8095(02)00076-5
Błaś M, Sobik M, Twarowski R (2008) Changes of cloud water chemical compositions in the Western Sudety Mountains, Poland. Atmos Res 87:224–231. https://doi.org/10.1016/j.atmosres.2007.11.004
Błaś M, Polkowska Ż, Sobik M, Klimaszewska K, Nowiński K, Namieśnik J (2010) Fog water chemical composition in different geographic regions of Poland. Atmos Res 95:455–469. https://doi.org/10.1016/j.atmosres.2009.11.008
Błaś M, Sobik M, Polkowska Ż, Cichała-Kamrowska K, Namieśnik J (2012) Water and chemical properties of hydrometeors over Central European mountains. Pure appl Geophys 5–6:1067–1081. https://doi.org/10.1007/s00024-011-0359-2
Bräuning A, De Ridder M, Zafirov N, García-González I, Dimitrov DP, Gärtner H (2016) Tree-ring features: indicators of extreme event impacts. IAWA J 37:131–206. https://doi.org/10.1163/22941932-20160131
Bytnerowicz A (1996) Physiological aspects of air pollution stress in forests. Phyton 36:15–22
Caporn SJM (2013) Ecophysiological responses of plants to air pollution. eLS. https://doi.org/10.1002/9780470015902.a0003206.pub2
Carlquist S (1988) Comparative wood anatomy. Systematic, ecological, and evolutionary aspects of dicotyledon wood. Springer, Berlin
Castagneri D, Fonti P, von Arx G, Carrer M (2017) How does climate influence xylem morphogenesis over the growing season? Insights from long-term intra-ring anatomy in Picea abies. Ann Bot 119:1011–1020. https://doi.org/10.1093/aob/mcw274
Černy M (1994) Monitoring of forests in the Czech Republic. In: Solon J, Roo-Zielihska E, Bytnerowicz A (eds) Climate and atmospheric deposition studies in forests, conference papers, vol 19, pp 87–100, 1GSO PAS, Warszawa
Cook ER, Holmes RL (1986) User’s manual for computer program ARSTAN. In: Holmes RL, Adams RK, Fritts HC (eds) Tree-ring chronologies of western North America: California, eastern Oregon and northern Great Basin, Chronology Ser. 6. University of Arizona, Tucson, pp 50–65
Danek M (2007) The influence of industry on scots pine stands in the south-eastern part of the Silesia-Krakow upland (Poland) on the basis of dendrochronological analysis. Water Air Soil Pollut 185:265–277. https://doi.org/10.1007/s11270-007-9448-3
de Kort I, Loeffen V, Baas P (1991) Ring width, density and wood anatomy of Douglas fir with different crown vitality. IAWA Bull 12:453–465. https://doi.org/10.1163/22941932-90000550
Denne MP (1988) Definition of latewood according to Mork (1928). IAWA Bull 10:59–62. https://doi.org/10.1163/22941932-90001112
Dore AJ, Choularton TW, Fowler D (1992) An improved wet deposition map of the United Kingdom incorporating the seeder-feeder effect over mountainous terrain. Atmos Environ 26A:1375–1381. https://doi.org/10.1016/0960-1686(92)90122-2
Dore AJ, Sobik M, Migała K (1999) Patterns of precipitation and pollutant deposition by rain and snow in the western Sudete Mountains, Poland. Atmos Environ 33:3301–3312. https://doi.org/10.1016/S1352-2310(98)00294-5
Dünisch O, Bauch J, Puls J, Müller M (1996) Biological and chemical wood properties of long-term polluted spruce (Picea abies [L.] Karst) at high-altitude stands of the Erzgebirge. Holzforschung 50:497–506
Elling W, Dittmar C, Pfaffelmoser K, Rötzer T (2009) Dendroecological assessment of the complex causes of decline and recovery of the growth of silver fir (Abies alba Mill.) in Southern Germany. For Ecol Manag 257:1175–1187. https://doi.org/10.1016/j.foreco.2008.10.014
Emberson L (2003) Air pollution impacts on crops and forests: an introduction. In: Emberson L, Ashmore M, Murray F (eds) Air pollution impacts on crops and forests: a global assessment. Imperial College Press, London, pp 3–29
EMEP (2018) Co-operative programme for monitoring and evaluation of the long range transmission of air pollutants in Europe. www.emep.int
Evert RF (2006) Esau’s plant anatomy. Meristems, cells, and tissues of the plant body—their structure, function, and development. John Wiley & Sons Inc., Hoboken
Evertsen JA, Mac Siurtain MP, Gardiner JJ (1986) The effects of industrial emission on wood quality in Norway spruce (Picea abies). IAWA Bull 7:399–404. https://doi.org/10.1163/22941932-90001010
Ferretti M, Innes JL, Jalkanen R, Saurer M, Schäffer J, Spiecker H, von Wilpert K (2002) Air pollution and environmental chemistry—what role for tree-ring studies? Dendrochronologia 20:159–174. https://doi.org/10.1078/1125-7865-00014
Fritts HC (1976) Tree rings and climate. Academic Press, New York
Godek M, Błaś M, Sobik M, Polkowska Ż, Cichała-Kamrowska K, Namieśnik J (2012) Environmental role of rime chemistry at selected mountain sites in Poland. Pure appl Geophys 169:1093–1106. https://doi.org/10.1007/s00024-011-0360-9
Grissino-Mayer HD, Holmes RI, Fritts HC (1996) COFECHA and ARSTAN—tree rings computer programs. University of Arizona, Tucson
Hauhs M, Wright RF (1988) Acid deposition: reversibility of soil and water acidification—a review. Air Pollution Report 11, CEC, Brussels
Hejnowicz Z (1971) Upward movement of the domain pattern in the cambium producing wavy grain in Picea excelsa. Acta Soc Bot Pol 40:499–512. https://doi.org/10.5586/asbp.1971.037
Hertzberg M, Aspeborg H, Schrader J, Andersson A, Erlandsson R, Nilsson J, Blomqvist K, Bhalerao R, Uhlén M, Teeri TT, Lundeberg J, Sundberg B, Nilsson P, Sandberg G (2001) A transcriptional roadmap to wood formation. PNAS 98:14732–14737. https://doi.org/10.1073/pnas.261293398
Igawa M, Kojima K, Yoshimoto O, Nanzai B (2015) Air pollutant deposition at declining forest sites of the Tanzawa Mountains, Japan. Atmos Res 151:93–100. https://doi.org/10.1016/j.atmosres.2014.03.017
Jadczyk P (2009) Natural effects of large-area forest decline in the Western Sudeten. Environ Prot Eng 35:49–56
Juknys R, Augustaitis A, Vencloviene J, Kliučius A, Vitas A, Bartkevičius E, Jurkonis N (2014) Dynamic response of tree growth to changing environmental pollution. Eur J For Res 133:713–724. https://doi.org/10.1007/s10342-013-0712-3
Klemm O, Wrzesinsky T (2007) Fog deposition fluxes of water and ions to a mountainous site in Central Europe. Tellus B 59:705–714. https://doi.org/10.1111/j.1600-0889.2007.00287.x
Kolář T, Čermák P, Oulehle F, Trnka M, Štěpánek P, Cudlín P, Hruška J, Büntgen U, Rybníček M (2015) Pollution control in the 1980s contributed to unprecedented spruce growth in the “Black Triangle”, the Czech-Polish border region. Sci Total Environ 538:703–711
Kopáček J, Veselý J (2005) Sulfur and nitrogen emissions in the Czech Republic and Slovakia from 1850 till 2000. Atmos Environ 39(12):2179–2188. https://doi.org/10.1016/j.atmosenv.2005.01.002
Kryza M, Błaś M, Dore AJ, Sobik M (2009) Modelling the concentration and deposition of air pollutants in Poland with the FRAME model. Dissertations of the Institute of Geography and Regional Development University of Wrocław 12, University of Wrocław
Kulhavy J, Klimo E (1998) Soil and nutrition status of forest stands under various site conditions of the Moravian–Silesian Beskids. Chemosphere 36(4–5):1113–1118
Kurczyńska EU, Dmuchowski W, Włoch W, Bytnerowicz A (1997) The influence of air pollution on needles and stems of Scots pine (Pinus silvestris L) trees. Environ Pollut 98:325–334. https://doi.org/10.1016/S0269-7491(97)00141-3
L’Hirondelle SJ, Addison PA (1985) Effects of SO2 on leaf conductance, xylem tension, fructose and sulphur levels of jack pine seedlings. Environ Pollut 39:373–386. https://doi.org/10.1016/0143-1471(85)90116-3
Lee EH, Beedlow PA, Waschmann RS, Tingey DT, Wickham C, Cline S, Bollman M, Carlile C (2016) Douglas-fir displays a range of growth responses to temperature, water, and Swiss needle cast in western Oregon, USA. Agric For Meteorol 221:176–188. https://doi.org/10.1016/j.agrformet.2016.02.009
Linzon SN (1971) Economic effects of sulphur dioxide on forest growth. J Air Pollut Control Assoc 21:81–86
Lloyd AH, Duffy PA, Mann DH (2013) Nonlinear responses of white spruce growth to climate variability in interior Alaska. Can J For Res 43:331–343. https://doi.org/10.1139/cjfr-2012-0372
Lovett GM (1984) Rates and mechanisms of cloud water deposition to a subalpine balsam fir forest. Atmos Environ 18:361–371. https://doi.org/10.1016/0004-6981(84)90110-0
Lukeš (2017) Meteorologická stanice Lysá hora. Retrieved October 15, 2018, from http://www.lysahora.cz/encyklopedie/objekty1.phtml?id=107685
Matschullat J, Maenhaut W, Zimmermann F, Fiebig J (2000) Aerosol and bulk deposition trends in the 1990′s. Eastern Erzgebirge; Central Europe. Atmos Environ 34:3213–3221. https://doi.org/10.1016/S1352-2310(99)00516-6
Mork E (1928) Die qualitat des Fichtenholzes unter besonderer Rücksichtnahme auf Schleif- und Papierholz. Der Papier Fabrikant 26:741–747
Myśkow E (2010) Procambium-cambium transition during vascular meristem development in Diospyros lotus. Botany 88:985–993. https://doi.org/10.1139/B10-070
Myśkow E, Zagórska-Marek B (2004) Ontogenetic development of storied ray pattern in cambium of Hippophae rhamnoides. Acta Soc Bot Pol 73:93–101. https://doi.org/10.5586/asbp.2004.014
Myśkow E, Zagórska-Marek B (2013) Dynamics of the ray pattern in cambium of Diospyros lotus. Dendrobiology 69:21–30
Nash TH, Fritts HC, Stokes MA (1975) A technique for examining non-climatic variation in widths of annual tree rings with special reference to air pollution. Tree-ring Bull 35:15–24
National Institute for public Health and Environment (2010) Effects of air pollution on European ecosystems, Technical report 2014, press release by Diego Fdez Sevilla. https://diegofdezsevilla.wordpress.com
Nilsson J, Grennfelt P (1988) Critical loads of sulphur and nitrogen. Nordic council of Ministers. Report 15, Copenhagen, Denmark, pp 8–57
Norton DA (1998) Impacts of tree coring on indigenous trees. Conservation Advisory Science Notes No. 186, Department of Conservation, Wellington
Nowosad M, Wereski S (2016) Warunki klimatyczne. In: Górecki A, Zemanek B (eds) Bieszczadzki Park Narodowy – 40 lat ochrony. Bieszczadzki Park Narodowy, Ustrzyki Górne, 31–38 (in Polish), http://serwisy.umcs.lublin.pl/marek.nowosad/publications%20and%20other%20papers%2F2016%20Warunki%20klimatyczne%20%5Bw%5D%20BPN%2040%20lat%20Ochrony.pdf
O’Brien TP, McCully ME (1981) The study of plant structure principles and selected methods. Termarcarphi Pty. Ltd., Melbourne
Oulehle F, Hleb R, Houška J, Šamonil P, Hofmeister J, Hruška J (2010) Anthropogenic acidification effects in primeval forests in the Transcarpathian Mts., western Ukraine. Sci Total Environ 408(4):856–864
Pahl S, Winkler P, Schneider T, Arends B, Schell D (1994) Deposition of trace substances via cloud interception on a coniferous forest at Kleiner Feldberg. J Atmos Chem 19:231–252. https://doi.org/10.1007/BF00696590
Regent Instruments Canada Inc. (2009) WinDENDRO for Tree-ring Analysis. Quebec. Canada. http://regent.qc.ca/assets/windendro_about.html
Rydval M, Wilson R (2012) The impact of industrial SO2 pollution on North Bohemia conifers. Water Air Soil Pollut 223:5727–5744. https://doi.org/10.1007/s11270-012-1310-6
Samusevich A, Zeidler A, Vejpustková M (2017) Influence of air pollution and extreme frost on wood cell parameters at mountain spruce stand (Picea abies (L.) Karst.) in the Ore Mountains. Wood Res 62:79–90
Schrader J, Nilsson J, Mellerowicz E, Berglund A, Nilsson P, Hertzberg M, Sandberg G (2004) A high-resolution transcript profile across the wood-forming meristem of poplar identifies potential regulators of cambial cell identity. Plant Cell 16:2278–2292. https://doi.org/10.1105/tpc.104.024190
Schweingruber FH (1988) Tree rings: basics and applications of dendrochronology. Kluwer Academic Publishing Group, Boston
Schweingruber FH (1996) Tree rings and environment. Dendroecology. Paul Haupt AG, Bern
Sensuła B, Opała M, Wilczyński S, Pawełczyk S (2015) Long- and short-term incremental response of Pinus sylvestris L. from industrial area nearby steelworks in Silesian Upland, Poland. Dendrochronologia 36:1–12. https://doi.org/10.1016/j.dendro.2015.08.001
Shigihara A, Matsumoto K, Sakurai N, Igawa M (2008) Leaching of cell wall components caused by acid deposition on fir needles and trees. Sci Total Environ 398:185–195. https://doi.org/10.1016/j.scitotenv.2008.03.007
Shimadera H, Kondo A, Lal Shrestha K, Kaga A, Inoue Y (2011) Annual sulfur deposition through fog, wet and dry deposition in the Kinki Region of Japan. Atmos Environ 45:6299–6308. https://doi.org/10.1016/j.atmosenv.2011.08.055
Sidor CG, Popa I, Vlad R, Cherubini P (2015) Different tree-ring responses of Norway spruce to air temperature across an altitudinal gradient in the Eastern Carpathians (Romania). Trees 29:985–997. https://doi.org/10.1007/s00468-015-1178-3
Skybova M (2006) Chemical composition of fog/cloud and rain water in the Beskydy Mountains—Czech Republic. Fresenius Environ Bull 15(5):448–451
Sobik M (1991) Precipitation and cloudwater acidity in the Karkonosze Mountains. Alpex Reg Bull 17:26–27
Song L, Kuang F, Skiba U, Zhu B, Liu X, Levy P, Dore A, Fowler D (2017) Bulk deposition of organic and inorganic nitrogen in southwest China from 2008 to 2013. Environ Pollut 227:157–166. https://doi.org/10.1016/j.envpol.2017.04.031
Szoege HM (2007) Sustainable development in Poland and Ukraine with special respect to rural areas. Manag Theory Stud Rural Bus Infrastruct Dev 8:129–137
Treml V, Ponocná T, Büntgen U (2012) Growth trends and temperature responses of treeline Norway spruce in the Czech-Polish Sudetes Mountains. Clim Res 55:91–103
Uggla C, Moritz T, Sandberg G, Sundberg B (1996) Auxin as a positional signal in pattern formation in plants. PNAS 93:9282–9286. https://doi.org/10.1073/pnas.93.17.9282
Vacek S, Bílek L, Schwarz O, Hejcmanová P, Mikeska M (2013) Effect of air pollution on the health status of spruce stands—a case study in the Krkonoše Mountains, Czech Republic. Mt Res Dev 33(1):40–50. https://doi.org/10.1659/MRD-JOURNAL-D-12-00028.1
Vestreng V, Myhre G, Fagerli H, Reis S, Tarrasón L (2007) Twenty-five years of continuous sulphur dioxide emission reduction in Europe. Atmos Chem Phys 7(13):3663–3681. https://doi.org/10.5194/acp-7-3663-2007
Weathers KC, Lovett GM, Likens GE (1995) Cloud deposition to a spruce forest edge. Atmos Environ 29:665–672. https://doi.org/10.1016/1352-2310(94)00317-E
Wilmking M, D’Arrigo R, Jacoby GC, Juday GP (2005) Increased temperature sensitivity and divergent growth trends in circumpolar boreal forests. Geophys Res Lett 32:L15715. https://doi.org/10.1029/2005GL023331
Wimmer R (2002) Wood anatomical features in tree-rings as indicators of environmental change. Dendrochronologia 20:21–36. https://doi.org/10.1078/1125-7865-00005
Wodzicki TJ (2001) Natural factors affecting wood structure. Wood Sci Technol 35:5–26. https://doi.org/10.1007/s002260100085
Yamaguchi T, Katata G, Noguchi I, Sakai S, Watanabe Y, Uematsu M, Furutani H (2015) Long-term observation of fog chemistry and estimation of fog water and nitrogen input via fog water deposition at a mountainous site in Hokkaido, Japan. Atmos Res 151:82–92. https://doi.org/10.1016/j.atmosres.2014.01.023
Zajączkowski S, Wodzicki TJ (1978) Auxin in plant morphogenesis—a model of regulation. Acta Soc Bot Pol 47:233–243. https://doi.org/10.5586/asbp.1978.021
Zar JH (1998) Biostatistical analysis, 4th edn. Prentice Hall, Upper Saddle River
Zawiła-Niedźwiedzki T (1994) Evaluation of forest state in threatened ecosystems with the use of satellite images and GIS. In: Proceedings of the Institute of Geodesy and Cartography, Warszawa
Acknowledgements
We thank the Editage service for English corrections. This study was supported by the research project no. 2012/15/B/ST10/00437 and 2015/17/B/ST10/03827 from the National Science Centre as well as partly by the University of Wrocław (Grant No. 0401/0423/17). We thank the two anonymous reviewers for extensive revision of our manuscript and useful comments allowing to improve a quality of the article.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Martina Meincken.
Rights and permissions
OpenAccess This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Myśkow, E., Błaś, M., Sobik, M. et al. The effect of pollutant fog deposition on the wood anatomy of subalpine Norway spruce. Eur J Forest Res 138, 187–201 (2019). https://doi.org/10.1007/s10342-018-01160-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-018-01160-4