
Abstract
Trophic relationships between Scorpaena porcus, S. notata and Chelidonichthys lucerna collected near natural gas platforms were investigated for the first time in central Adriatic Sea from July 2005 to May 2006. Sampling was repeated at control sites on soft bottom where, however, only C. lucerna occurred. All of the three predators showed diet specialization for crustaceans, with high overlap for angular crab Goneplax rhomboides. The widest prey variety, consisting mainly of epibenthic and infaunal organisms, was detected in C. lucerna, whose feeding behaviour resulted not to be affected by the presence of platforms. Burrowing shrimp Alpheus glaber, hairy crab Pilumnus hirtellus, bivalve Corbula gibba, and European anchovy Engraulis encrasicolus discriminated S. porcus diet, while thalassinid Jaxea nocturna discriminated S. notata diet. The occurrence of E. encrasicolus as natural prey of S. porcus is discussed. Overall, predators like scorpaenids, which live in strict association with platforms in the study area, seem to rely on soft bottom prey items rather than exploiting resources from the platforms, thus competing with soft-bottom predators (i.e. triglids) for the same resources.
Similar content being viewed by others

Introduction
Oil and natural gas platforms placed on soft-sediment bottoms act as attractors for many species, thus increasing biodiversity in the surrounding marine environment (Fabi et al. 2004; Stanley and Wilson 1997; Wolfson et al. 1979). They act as artificial reefs that provide new attachment sites for benthic organisms and vertical relief attractive to fishes, inducing the development of new organism assemblages (Stachowitsch et al. 2002; Terlizzi et al. 2008). In particular, fish assemblages associated with these offshore structures encompass both soft-bottom dwelling species, which are also found in the original substrate, and reef dwelling species, which would hardly (if not at all) be represented without these permanent structures. A pelagic component in a varying level of abundance has also been recorded near offshore oil and natural gas platforms (Franks 2000; Neira 2005). This pattern is confirmed for many platforms that were investigated in different areas of the world, such as southern California (Love et al. 2000; Martin and Lowe 2010), Gulf of Mexico (Hastings et al. 1976; Stanley and Wilson 1997), African western coast (Gerlotto et al. 1989), North Sea (Jørgensen et al. 2002), and the Mediterranean (Andaloro et al. 2011; Consoli et al. 2007; Fabi et al. 2004), although species composition varies according to the specific area concerned.
Although the effects of platforms on biodiversity, particularly as regards fish assemblages, have been investigated, their role on trophic relationships of their associated organisms have so far received relatively little attention. In this regard, some useful information can be drawn from specific studies carried out near artificial reefs, which are hard substrates lying on soft bottoms like oil or natural gas platforms. In general terms, only few fish species seem to be attracted by artificial reefs for food (Steimle and Ogren 1982), showing a different degree of dependence from reef resources, where reef dwelling fishes are the major exploiters. In particular, brown meagre Sciaena umbra in the Adriatic (Fabi et al. 2006) and white seabream Diplodus sargus in Portuguese waters (Leitão et al. 2007) better than other artificial reef fish species exploit reef resources. Other species, which are not strictly associated with hard bottoms, such as shi drum Umbrina cirrosa, annular seabream Diplodus annularis and striped seabream Lithognathus mormyrus, partially rely on reef resources for their diet (Fabi et al. 1998, 2006). In the Atlantic, Hueckel and Stayton (1982) detected a direct correlation between the diet of artificial reef fish species (i.e. striped seaperch Embiotoca lateralis, pile perch Rhacochilus vacca and quillback rockfish Sebastes maliger) and the organisms growing on artificial reefs. Said correlation is stronger for medium- and large-sized predators, while smaller-sized predators are more likely to use artificial reefs as a shelter. Conversely, in Mediterranean artificial reefs, scorpaenid species (i.e. black scorpionfish Scorpaena porcus and small red scorpionfish S. notata) rarely or do not feed on reef species but, rather, explore the surrounding sediment for food (Relini et al. 2002).
In most cases, the greatest biomass of reef dwelling species living near platform structures is made up of rockfish from the genus Sebastes at various life stages (Love et al. 2000). In the case of platforms in the Mediterranean, these mainly consist of scorpaenid species from the genus Scorpaena (Fabi et al. 2004) that are possibly attracted by shelter or food availability. Scorpaenids are known to be highly selective predators and principally feed on crustacean decapods (Harmelin-Vivien et al. 1989; Follesa et al. 2004; Morte et al. 2001), which are also commonly preyed on by soft-bottom scorpaeniforms, for instance triglids (Colloca et al. 1994; Morte et al. 1997; Nouvel 1950). Both scorpaenids and triglids are highly abundant near platforms in the Adriatic Sea (Andaloro et al. unpublished data). In particular, S. porcus and S. notata from the Scorpaenidae family and Chelidonichthys lucerna from the Triglidae family are the most common species and possibly compete for the same food resources. We investigated trophic relationships between these three species living at and near natural gas platforms.
Materials and methods
Study site
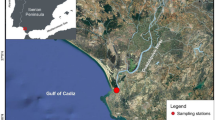
From July 2005 to May 2006, four seasonal fishery surveys (July, October, January, May) were carried out by commercial trawl and bottom gillnet around two cluster gas platforms in the Abruzzi coastal waters, central Adriatic Sea (Mediterranean). These two platforms, named Fratello Cluster and Fratello Nord, are placed on a sand-muddy bottom at 19 and 23 m depth, respectively (Fig. 1). Fish were collected near both platforms and at control sites located 5 km approximately from the platforms but at the same bathymetry.

Map of the study area with indication of the platforms and control site investigated
Bottom gillnet (416 m long and 3.40 m deep, with a 34-mm stretched mesh size) was set within a 50 m radius from the platforms and at control sites; it was lowered at dusk and hauled at dawn. Trawl was hauled for 20 min at an average speed of 2.8 knots, covering a distance of approximately 1,700 m; hauls began at sunrise and continued up until late morning both near the platforms and at control sites.
Each specimen was measured to the nearest 1 mm total length (L T) and weighed to the nearest 0.1 g. Fish were eviscerated and their stomachs preserved in 70% ethanol solution. Prey items were sorted and identified to the lowest possible taxonomic level, counted and weighed to the nearest 0.1 mg.
Data analyses
The number of empty stomachs for each species was recorded and used to calculate the vacuity coefficient Cv% (=number of empty stomachs/total number of analysed stomachs × 100).
The importance of the different prey categories was assessed by calculating the frequency of occurrence (%F; percentage of stomachs containing a prey item or category), abundance (%N; number of specimens of a specific prey item or category as a percentage of the total number of prey items found) and mass (%W; mass of a prey item or category as a percentage of the total mass of prey ingested). These values were used to calculate the index of relative importance (IRI) for each category (Hyslop 1980) using mass instead of volume (Hacunda 1981):
This index was expressed in percentage points (%IRI; Cortés 1997).
Diet breadth was calculated using Levin’s standardized index (Krebs 1989),
where p ij is the proportion of predator i diet made up of prey j, and n is the number of prey categories. This index ranges from 0 to 1; low values indicate a specialist predator with a diet dominated by few prey items, while high values indicate a generalist predator (Krebs 1989).
Niche overlap was calculated separately using Morisita’s index (1959), which is considered the most sound index when the number of prey individuals is known (Smith and Zaret 1982). This index (C) is calculated as
where: p ij, p ik = proportion of prey resource i as to total prey resources exploited by species j and species k, n ij n ik = number of individuals of species j and k that exploit resource i; N j , N k = total number of sampled individuals from each species. C ranges from 0 (when predators have no prey species in common) to 1 (when prey species composition is identical among predators).
A non-parametric multivariate analysis of variance (PERMANOVA) (Anderson et al. 2008) was performed on prey abundance data to detect differences between sampling periods (two-factor analysis: predators × periods) and between predators (single-factor analysis), keeping separate data of C. lucerna caught near the platforms from those caught at control sites. Data were transformed to Log(x + 1); the analysis is based on Gower distances excluding double zeros and using 999 permutations. Pair-wise a posteriori comparisons were computed after significant differences among factor levels were detected.
PERMANOVA was also performed to detect differences between five predator size classes (two-factor analysis: predators × size classes) as follows: size class I ≤ 140 mm, size class II = 141–180 mm L T, size class III = 181–220 mm L T, size class IV = 221–260 mm L T, size class V > 260 mm L T. Such intervals were identified taking into account the size at which each species undergoes sexual maturity: (1) S. notata undergoes maturity at about 10–14 cm L T (Scarcella et al. 2011b); (2) C. lucerna starts maturing at sizes >180 mm L T (Montanini et al. 2008; Morte et al. 1997). The specimens of S. porcus collected in our study, except for only one specimen of 90 mm L T, exceeded the size at first maturity reported in literature (i.e. 108 mm L T for females and 85 mm L T for males) (Bradai and Bouain 1991), therefore, the size classes for this species are represented only by adult specimens. This statistical analysis was integrated with pair-wise a posteriori comparisons between size classes within levels of each factor.
Correspondence Discriminant Analysis (CDA) was computed on prey abundance data to examine the interactive effects between predators. Initially developed to examine ecological data, this analysis is appropriate for comparing individuals belonging to several distinct groups (Chessel and Thioulouse 1996). As CDA can be used on contingency tables and operates on correspondences between factors, it is particularly suitable for analysing unbalanced groups. It is a highly effective tool because it combines the advantages of standard Correspondence Analysis and Discriminant Analysis (Perrière et al. 1996). Monte Carlo test (RV) consisting of 999 repeated random permutations of rows between pre-defined groups was performed to assess the significance of the discrimination.
CDA and Monte Carlo tests were repeated on the specimens caught by gillnet so as to exclude any possible biased results owed to the use of different gears (trawl and gillnet) for the collection of the specimens. CDA and Monte Carlo test were computed with modules included in ADE4 (a package of the R software) (Dray and Dufour 2007).
Results
A total of 572 fishes were analysed for stomach content, length and vacuity coefficient (Table 1). Black scorpionfish Scorpaena porcus and small red scorpionfish S. notata were only caught near the platforms, almost exclusively by gillnet (only three specimens of S. porcus and one of S. notata were caught by trawl), while tub gurnard Chelidonichthys lucerna was collected both near the platforms (N = 219) and at control (N = 168) sites via both gears: 120 and 267 specimens were collected by trawl and by gillnet, respectively.
About 32.3% of the stomachs analysed were found empty; scorpaenids showed the highest vacuity coefficient. C. lucerna showed a different Cv% according to the gear used (Cv% = 25.5 for individuals caught by gillnet and Cv% = 6.7 for individuals caught by trawl).
The analysis of prey items led to the identification of 53 taxa belonging to five principal taxonomic groups; 15 taxa were detected in S. notata, 9 in S. porcus and 51 in C. lucerna. Crustaceans were dominant in all three predators examined according to all numeric indicators, scoring values above 82%. Teleosts were the second most important prey group, with higher values recorded in C. lucerna than in the two scorpaenids and %F, %N, %W and % IRI values are reported in Table 2.
Angular crab Goneplax rhomboides was the most important prey item for all three predators. The second most important prey organism was pistol shrimp Alpheus glaber which was more represented in S. porcus stomach contents compared to S. notata and C. lucerna.
Levin’s standardized index calculated for prey category numerical abundance was 0.25 for S. notata, 0.12 for S. porcus, and 0.03 for C. lucerna.
Morisita’s index values of niche overlap between pairs of predators were as follows: C notata-porcus = 0.69, C notata-lucerna = 0.66, C porcus-lucerna = 0.78.
PERMANOVA performed on prey abundance produced no significant differences between sampling periods (F 3,372 = 1.1366 n.s.) or with regard to the predator × period interaction (F 8,372 = 1.3203 n.s.), while it showed highly significant differences among predators (F 3,372 = 2.6815, P < 0.01). Such differences are confirmed by single-factor PERMANOVA (F 3,383 = 3.4993, P < 0.01) with pair-wise a posteriori comparisons resulting as follows: S. porcus ≠ S. notata≠ (C. lucerna platform = C. lucerna control).
No significant differences were detected between predator size classes (F 4,376 = 1.3447 n.s.), nor in the predator × size class interaction (F 4,376 = 0.8966 n.s.). Pair-wise a posteriori comparisons revealed that the size class V, only present in C. lucerna, was significantly different (P < 0.05) from all the other size classes of all predators.
The discrimination operated by CDA produced significant results (RV = 0.15, P < 0.05) that are visualized in a composed plot (Fig. 2). As shown in the correlation circle (Fig. 2a), some prey items are more important than others in function of their major distance from origin and major proximity to x and y axes. For a complete information, prey items in the first quadrant of the correlation circle (rectangle) are expanded in Fig. 2b. This analysis highlighted an opposite trend between S. porcus and S. notata, while C. lucerna holds an intermediate position between the two scorpaenids (Fig. 2c). S. porcus diet is discriminated by prey organisms that are parallel to x axes, principally those on the left of the correlation circle; S. notata diet is discriminated by prey organisms that are parallel to y axes, in particular those on the lower part of the correlation circle. Prey items that C. lucerna feeds on are mainly concentrated around the centre of the correlation circle. Figure 2d shows the gravity centres of the ellipses reported in Fig. 2c, corresponding to the exact position of each predator to be compared with the position of prey items in Fig. 2a.

Correspondence discriminant analysis (CDA) computed on prey abundance data for Scorpaena porcus, S. notata and Chelidonichthys lucerna. a Correlation circle between discriminant variables (prey items) and discriminant functions, b expansion of the rectangle evidenced in (a), c projection of the stomach contents with ellipses and gravity centres of predators, d plot of gravity centres of predators
CDA performed on the samples collected by gillnet produced significant results (RV = 0.015, P < 0.05): the trend described above was confirmed (i.e. orthogonality between S. porcus and S. notata, with C. lucerna at an intermediate level between them as shown in Fig. 3b), with the same discriminated prey organisms (Fig. 3a), except for Engraulis encrasicolus.

Correspondence discriminant analysis (CDA) computed on prey abundance data for Scorpaena porcus, S. notata and Chelidonichthys lucerna collected by gillnet. a Correlation circle between discriminant variables (prey items) and discriminant functions, b projection of the stomach contents with ellipses and gravity centres of predators
Discussion
Oil and natural gas platforms may represent an essential habitat for some organisms (Helvey 2002), in particular for reef dwelling species that need hard substrates to survive. In the western Adriatic Sea, these species are patchily distributed since the seabed principally consists of wide soft bottoms with rare areas of rocky grounds and various inshore artificial substrates. In this context, platforms that are located on sandy grounds far from rocky areas may be considered like ‘oases in the desert’, which provide additional habitat to species that would not otherwise settle there. This is evidenced by the frequent occurrence of scorpaenids near many Adriatic platforms, and their scarcity or absence from the surrounding areas (Andaloro et al. unpublished data; Fabi et al. 2004). This trend is further confirmed by this study as scorpaenids were almost exclusively caught near the platforms.
Stomach content analysis of Chelidonichthys lucerna, Scorpaena porcus and S. notata highlighted some differences between these three predators, which are not related to the different sampling periods based on multivariate analysis results. Scorpaenids showed the highest vacuity index. As they were caught almost exclusively by gillnet, this result is possibly ascribable to the fishing gear technique used, which requires the net to be set for long time. As these species are night predators (Harmelin-Vivien et al. 1989), several of them might have been entangled before prey ingestion, or digestion process might have concluded during their permanence in the net before drawing up. This result is also supported by the higher vacuity index recorded for C. lucerna specimens caught by gillnet (Cv% = 25.5) as compared to results obtained for trawled specimens (Cv% = 6.7).
Diet breadth analysis of C. lucerna, S. porcus and S. notata has evidenced that all three predators show specialist feeding behaviours, as confirmed by Levin’s index very low values. Diet analysis indicates that they mostly feed on crustaceans, while the other prey groups detected (i.e. teleosts, molluscs, salpids, sipunculans) only play a marginal role in their diet.
As highlighted by %IRI values, all predators massively feed on angular crab Goneplax rhomboides, a very common species on Adriatic soft bottoms (Scarcella et al. 2007). A comparison between the diets of the three predators has evidenced the widest prey variety in C. lucerna; said array is mainly made up of epibenthic and infaunal organisms encompassing a variety of crustaceans, molluscs and fish that are typical of soft bottoms. Said feeding behaviour is in line with what reported in literature for this species in other areas (Collignon and Aloncle 1960; Colloca et al. 1994; Costa 1988; Morte et al. 1997; Stagioni et al. 2011), although prey composition may vary depending on local resource availability.
As artificial substrates deployed on soft bottoms alter biodiversity, favouring either the settlement of hard bottom species on the structures themselves or the development of deposit and suspension feeders in the surrounding sediment (Fabi et al. 2002), it might be expected that the type of prey differs when predators live near or far from the structures. In our study, the similarity between the diet of tub gurnards caught near the platforms and of those caught at control sites suggests that artificial structures do not affect this predator’s prey choice, since it consistently feeds on soft-bottom organisms whose occurrence in the study area does not depend on the presence of hard substrates.
Chelidonichthys lucerna diet strongly overlaps with S. porcus and S. notata to such an extent that the latter’s diets are subsets of the former’s diet in terms of prey array. However, some specific prey organisms characterize the diet of one or of the other predator either because they are exclusively found in only one of the three predators or because their abundance level is different. In particular, S. porcus, which shows a diet based on few prey items, feeds with relatively high frequency on burrowing shrimp Alpheus glaber and, to a lesser extent, on hairy crab Pilumnus hirtellus, on bivalve Corbula gibba, and on European anchovy Engraulis encrasicolus. These prey species discriminate black scorpionfish diet from the other two predators, as highlighted by CDA (Fig. 2). While A. glaber and C. gibba are mud-loving species, commonly found in Adriatic soft bottoms, P. hirtellus is more commonly found on rocky substrates and on banks of mussels (Stevcic 1990); and its occurrence in the area might be favoured by the presence of platform structures (Manoukian et al. 2010). In natural rocky and rocky-sandy substrates, this species represents one of the most important organisms that black scorpionfish preys on (Pallaoro and Jardas 1991). The occurrence of E. encrasicolus, which is one of the most common pelagic species in the study area, in S. porcus stomach contents is not new to our knowledge. Demirhan and Can (2009) reported that this species are among the organisms that Black Sea S. porcus commonly preys on. This would indicate that black scorpionfish exploits not only benthic species, but also relies on water column organisms for its diet. This hypothesis, however, needs further confirmation since CDA performed exclusively on gillnet samples (see Fig. 3) did not discriminate European anchovy among black scorpionfish prey organisms. This might indicate that the results were possibly biased by the sampling gear used: in fact, European anchovies found in trawled specimens’ stomachs might have been eaten inside the trawl net. Overall feeding behaviours highlighted for S. porcus in this study are in line with what reported in literature in other areas, where this predator diet is mainly based on crustaceans and, only secondarily, on fishes, rarely on molluscs and polychaetes (Arculeo et al. 1989; Başçınar and Sağlam 2009; Bradai and Bouain 1990; Carpentieri et al. 2001; Demirhan and Can 2009; Follesa et al. 2004; Harmelin-Vivien et al. 1989; Morte et al. 2001; Pallaoro and Jardas 1991); nonetheless, it must be noted that prey composition varies according to the area examined. From our results, a similar feeding behaviour, mainly based on crustaceans, is also detected in S. notata. Despite the lower number of full stomachs examined compared to S. porcus, S. notata showed a more diversified prey array (15 prey taxa in the latter vs. 9 found in the former). The most important prey organisms for S. notata are G. rhomboides, followed by crangonid Philocheras bispinosus, thalassinid Jaxea nocturna and processid Processa macrophthalma. According to CDA, J. nocturna is the main prey item discriminating small red scorpionfish diet; this analysis also highlights a small group of prey organisms (i.e. gammarideans, Callianassa sp., Lesueurigobius juv., processids, crangonids) that are also preyed on by C. lucerna but to a lesser extent, while they lack in S. porcus diet. These results confirm that S. notata feeds on both epibenthic organisms and burrower ones, many of which are known to be nocturnal (Manning and Chace 1971; Pervesler and Dworschak 1985). The occurrence of some prey organisms like J. nocturna and P. macrophthalma, reported for the first time in this study for S. notata diet, possibly depends on their large availability in the study area.
The results of the analyses by size between predators suggest that there is no sharp divergence in prey abundance between size classes, except for size class V (L T > 260 mm)—only represented in C. lucerna—which resulted to be significantly different from all of the other size classes examined. Diet variation by size in C. lucerna was also reported by Colloca et al. (1994) in the Tyrrhenian Sea and by Stagioni et al. (2011) in the Adriatic Sea, albeit at smaller sizes (i.e. 14–16 cm standard length and 180 mm L T, respectively), which are possibly ascribable to the threshold from a juvenile to an adult stage. Such discrepancy might be related to the fact that oil and gas platforms represent particular environments where biological parameters may be altered, as testified by the positive growth performance showed by scorpaenids associated with northern Adriatic platforms as compared to those living in natural environments (Scarcella et al. 2011a).
Unlike summer decreases in feeding activity reported for these three species in natural environments, which are possibly related to reproductive events (Bradai and Bouain 1990; Pallaoro and Jardas 1991; Morte et al. 2001; Stagioni et al. 2011), our results showed no significant temporal variations in prey abundance, thus suggesting a constant resource exploitation by the three predators throughout the year.
Overall, predators like scorpaenids, which live in strict association with platforms in the study area, seem to rely on soft-bottom prey items rather than exploiting resources from the platforms. A similar behaviour was noticed in scorpaenids that live in association with artificial reefs, which prefer to explore the surrounding grounds for nourishment and feed very little on reef species (Relini et al. 2002). Therefore, platform-induced occurrence of scorpaenids on natural soft bottoms entails an overlap of the trophic niche between scorpaenids and the predators that are typical of soft bottoms, such as tub gurnard, leading to a competition for the same resources. Although the three predators examined massively exploit the same prey organisms (i.e. G. rhomboides), most probably because of their large availability in the area, they still show distinctive elements that characterize their diets, which let us hypothesize a certain degree of prey partitioning.
References
Andaloro F, Castriota L, Ferraro M, Romeo T, Sarà G, Consoli P (2011) Evaluating fish assemblages associated with gas platforms: evidence from a visual census technique and experimental fishing surveys. Cienc Mar 37(1):1–9
Anderson MJ, Gorley RN, Clarke KR (2008) PERMANOVA+ for PRIMER: guide to software and statistical methods. PRIMER-E, Plymouth
Arculeo M, Froglia C, Riggio S (1989) Considerazioni sull’alimentazione di alcune specie ittiche dei fondali infralitorali del Golfo di Palermo. Oebalia NS 15(1):57–65
Başçınar NS, Sağlam H (2009) Feeding habits of black scorpionfish Scorpaena porcus, in the South-Eastern Black Sea. Turk J Fish Aquat Sci 9:99–103
Bradai MN, Bouain A (1990) Régime alimentaire de Scorpaena porcus et de S. scrofa (Teleostei, Scorpaenidae) du Golfe De Gabes, Tunisie. Cybium 14(3):207–216
Bradai MN, Bouain A (1991) Reproduction de Scorpaena porcus (Linnaeus, 1758) et de Scorpaena scrofa (Linnaeus, 1758) (Pisces, Scorpaenidae) du Golfe de Gabes. Oebalia 17:167–180
Carpentieri P, Colloca F, Belluscio A, Ardizzone GD (2001) Preliminary notes on feeding habits of Scorpaena porcus (Linnaeus, 1758) in the central Tyrrhenian Sea. Biol Mar Mediterr 8(1):699–703
Chessel D, Thioulouse J (1996) L’analyse discriminante des correspondances. Documentation Thématique ADE-4, vol 5, sect. 6. Université Claude Bernard, Lyon 1, Lyon
Collignon J, Aloncle H (1960) Le régime alimentaire de quelques poissons benthiques des côtes marocaines. Bull Inst Pêches Marit Maroc 5:1–13
Colloca F, Ardizzone GD, Gravina MF (1994) Trophic ecology of gurnards (Pisces: Triglidae) in the Central Mediterranean Sea). Mar Life 4(2):45–57
Consoli P, Azzurro E, Sarà G, Ferraro M, Andaloro F (2007) Fish diversity associated with gas platforms: evaluation of two underwater visual census techniques. Cienc Mar 33(2):121–132
Cortés E (1997) A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes. Can J Fish Aquat Sci 54:726–738
Costa MJ (1988) Ecologie alimentaire de quelques poisons de l’estuaire du Tage. Cybium 12:301–320
Demirhan SA, Can MF (2009) Age, growth and food composition of Scorpaena porcus (Linneaus, 1758) in the southeastern Black Sea. J Appl Ichthyol 25:215–218
Dray S, Dufour AB (2007) The ade4 package: implementing the duality diagram for ecologists. J Stat Softw 22(4):1–20
Fabi G, Camilletti E, Ciccotti E, Luccarini F, Lucchetti A, Panfili M, Solustri C (1998) Ruolo trofico della barriera artificiale di Cesano-Senigallia nei confronti di alcune specie ittiche. Biol Mar Mediterr 5(3):1812–1821
Fabi G, Luccarini F, Panfili M, Solustri C, Spagnolo A (2002) Effects of an artificial reef on the surrounding soft-bottom community (central Adriatic Sea). ICES J Mar Sci 59:S343–S349
Fabi G, Grati F, Puletti M, Scarcella G (2004) Effects on fish community induced by installation of two gas platforms in the Adriatic Sea. Mar Ecol Prog Ser 273:187–197
Fabi G, Manoukian S, Spagnolo A (2006) Feeding behavior of three common fishes at an artificial reef in the Northern Adriatic Sea. Bull Mar Sci 78(1):39–56
Follesa MC, Cabiddu S, Sabatini A, Cau A (2004) Relazioni trofiche nelle specie di Scorpaena (Linneo, 1758) del Mediterraneo centro-occidentale. Biol Mar Mediterr 11(2):586–591
Franks JS (2000) A review: pelagic fishes at petroleum platforms in the northern Gulf of Mexico; diversity, interrelationships, and perspectives. In: Le Gall J-Y, Cayré P, Taquet M (eds) Pêche thonière et dispositifs de concentration de poissons. Actes de colloques 28. Ifremer, Plouzané, pp 502–515
Gerlotto F, Bercy C, Bordeau B (1989) Echo integration survey around offshore oil extraction platforms off Cameroon: observations of the repulsive effect on fish of some artificially emitted sounds. Proc Inst Acoust 19:79–88
Hacunda JS (1981) Trophic relationships among demersal fishes in a coastal area of the Gulf of Maine. Fish Bull 79:775–788
Harmelin-Vivien ML, Kaim-Malka RA, Ledoyer M, Jacob-Abraham SS (1989) Food partitioning among scorpaenid fishes in Mediterranean seagrass beds. J Fish Biol 34:715–734
Hastings RW, Ogren LH, Mabry MT (1976) Observations on the fish fauna associated with offshore platforms in the north-eastern Gulf of Mexico. Fish Bull US Nat Mar Fish Serv 74:387–401
Helvey M (2002) Are southern California oil and gas platforms essential fish habitat? ICES J Mar Sci 59:S266–S271
Hueckel GJ, Stayton RL (1982) Fish foraging on an artificial reef in Puget Sound, Washington. Mar Fish Rev 44(6–7):38–44
Hyslop EJ (1980) Stomach content analysis—a review of methods and their application. J Fish Biol 17:411–429. doi:10.1111/j.1095-8649.1980.tb02775.x
Jørgensen T, Løkkeborg S, Soldal AV (2002) Residence of fish in the vicinity of a decommissioned oil platform in the North Sea. ICES J Mar Sci 59:S266–S271
Krebs CJ (1989) Ecological methodology. Harper Collins Publishers, New York
Leitão F, Santos MN, Monteiro CC (2007) Contribution of artificial reefs to the diet of the white sea bream (Diplodus sargus). ICES J Mar Sci 64:473–478
Love MS, Caselle JE, Snook L (2000) Fish assemblages around seven oil platforms in the Santa Barbara Channel area. Fish Bull 98:96–117
Manning RB, Chace FA Jr (1971) Shrimps of the family Processidae from the northwestern Atlantic Ocean (Crustacea: Decapoda: Caridea). Smithson Contrib Zool 89:1–41
Manoukian S, Spagnolo A, Scarcella G, Punzo E, Angelini R, Fabi G (2010) Effects of two offshore gas platforms on soft-bottom benthic communities (northwestern Adriatic Sea, Italy). Mar Environ Res 70:402–410
Martin CJB, Lowe CG (2010) Assemblage structure of fish at offshore petroleum platforms on the San Pedro Shelf of southern California. Mar Coast Fish Dyn Manag Ecosyst Sci 2:180–194
Montanini S, Stagioni M, Vallisneri M (2008) Note sulla biologia di Chelidonichthys lucernus (Teleostei: Triglidae) nell’alto-medio Adriatico. Biol Mar Mediterr 15(1):340–341
Morisita M (1959) Measuring of interspecific association and similarity between communities. Mem Fac Sci Kyushu Univ Ser E (Biology) 3:65–80
Morte S, Redon MJ, Sanz-Brau A (1997) Trophic relationships between two gurnards Trigla lucerna and Aspitrigla obscura from the western Mediterranean. J Mar Biol Ass UK 77:527–537
Morte S, Redon MJ, Sanz-Brau A (2001) Diet of Scorpaena porcus and Scorpaena notata (Pisces: Scorpaenidae) in the western Mediterranean. Cah Biol Mar 42:333–344
Neira FJ (2005) Summer and winter plankton fish assemblages around offshore oil and gas platforms in south-eastern Australia. Estuar Coast Shelf Sci 63:589–604
Nouvel H (1950) Recherches sur la nourriture de quelques Trigles du Golfe de Gascogne au large d’Arcachon. Bull Inst Océanogr Monaco 964:1–12
Pallaoro A, Jardas I (1991) Food and feeding habits of black scorpionfish (Scorpaena porcus L. 1758) (Pisces, Scorpaenidae) along the Adriatic coast. Acta Adriat 32(2):885–898
Perrière G, Lobry JR, Thioulouse J (1996) Correspondence discriminant analysis: a multivariate method for comparing classes of protein and nucleic acid sequences. Comput Appl Biosci 12(6):519–524
Pervesler P, Dworschak PC (1985) Burrows of Jaxea nocturna Nardo in the Gulf of Trieste. Senckenb Marit 17(1–3):33–53
Relini G, Relini M, Torchia G, De Angelis G (2002) Trophic relationships between fishes and an artificial reef. ICES J Mar Sci 59:S36–S42
Scarcella G, Fabi G, Grati G (2007) Rapido trawl fishery in the north-central Adriatic Sea. Rapp Comm int Mer Médit 38:591
Scarcella G, Grati F, Polidori P, Bolognini L, Domenichetti F, Fabi G (2011a) Comparison of growth rates estimated by otolith reading of Scorpaena porcus and Scorpaena notata caught at artificial and natural reefs of the northern Adriatic Sea. Braz J Ocean 59(special issue CARAH):33–42
Scarcella G, La Mesa M, Grati G, Polidori P (2011b) Age and growth of the small red scorpionfish, Scorpaena notata Rafinesque, 1810, based on whole and sectioned otolith readings. Environ Biol Fish 91:369–378
Smith EP, Zaret TM (1982) Bias in estimating niche overlap. Ecology 63:1248–1253
Stachowitsch M, Kikinger R, Herler J, Zolda P, Geutebruck E (2002) Offshore oil platforms and fouling communities in the southern Arabian Gulf (Abu Dhabi). Mar Poll Bull 44(9):853–860
Stagioni M, Montanini S, Vallisneri M (2011) Feeding of tub gurnard Chelidonichthys lucerna (Scorpaeniformes: Triglidae) in the north-east Mediterranean. J Mar Biol Ass UK. doi:10.1017/s0025315411000671
Stanley DR, Wilson CA (1997) Seasonal and spatial variation in the abundance and size distribution of fishes associated with petroleum platform in the northern Gulf of Mexico. Can J Fish Aquat Sci 54:1166–1176
Steimle FW, Ogren L (1982) Food of fish collected on artificial reefs in the New York Bight and off Charleston, South Carolina. Mar Fish Rev 44(6–7):49–52
Stevcic Z (1990) Check-list of the Adriatic decapod Crustacea. Acta Adriat 31(1/2):183–274
Terlizzi A, Bevilacqua S, Scuderi D, Fiorentino D, Guarnieri G, Giangrande A, Licciano M, Felline S, Fraschetti S (2008) Effects of offshore platforms on soft-bottom macro-benthic assemblages: a case study in a Mediterranean gas field. Mar Poll Bull 56:1303–1309
Wolfson A, Van Blaricom G, Davis N, Lewbel GS (1979) The marine life of an offshore oil platform. Mar Ecol Prog Ser 1:81–89
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Arne Malzahn.
Rights and permissions
About this article
Cite this article
Castriota, L., Falautano, M., Finoia, M.G. et al. Trophic relationships among scorpaeniform fishes associated with gas platforms. Helgol Mar Res 66, 401–411 (2012). https://doi.org/10.1007/s10152-011-0281-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10152-011-0281-0