
Abstract
There are two kinds of neurotransmissions that occur in brain. One is neuron to neuron at synapses, and the other is neuron to glia via extracellular fluid (ECF), both of which are important for maintenance of proper neuronal functioning. For neuron to neuron communications, several potent amino acid neurotransmitters are used within the confines of synaptic space. However, their presence at elevated concentrations in extra-synaptic space could be detrimental to well organized neuronal functioning. The significance of the synthesis and release of N-acetylaspartylglutamate (NAAG) by neurons has long been a puzzle since glutamate (Glu) itself is the “key” that can interact with all Glu receptors on membranes of all cells. Nonetheless, neurons synthesize this acetylated dipeptide, which cannot be catabolized by neurons, and release it to ECF where its specific physiological target is the Glu metabotropic receptor 3 on the surface of astrocytes. Since Glu is excitotoxic at elevated concentrations, it is proposed that formation and release of NAAG by neurons allows large quantities of Glu to be transported in ECF without the risk of injurious excitotoxic effects. The metabolic mechanism used by neurons is a key–lock system to detoxify Glu during its intercellular transit. This is accomplished by first synthesizing N-acetylaspartate (NAA), and then joining this molecule via a peptide bond to Glu. In this paper, a hypothesis is presented that neurons synthesize a variety of relatively nontoxic peptides and peptide derivatives, including NAA, NAAG, homocarnosine (γ-aminobutyrylhistidine) and carnosine (β-alanylhistidine) from potent excitatory and inhibitory amino acids for the purpose of releasing them to ECF to function as cell-specific neuron-to-glia neurotransmitters.


Similar content being viewed by others

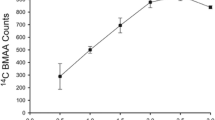
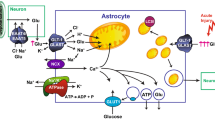
Abbreviations
- Ac:
-
Acetate
- AcCoA:
-
Acetylcoenzyme A
- Asp:
-
Aspartate
- ASPA:
-
Aspartoacylase
- ATP:
-
Adenosine triphosphate
- β-ala:
-
β-Alanine
- CD:
-
Canavan disease
- Carn:
-
Carnosine
- CSF:
-
Cerebrospinal fluid
- ECF:
-
Extracellular fluid
- ECS:
-
Extracellular space
- GABA:
-
γ-Aminobutyric acid
- Glu:
-
Glutamate
- GRM3:
-
Metabotropic Glu receptor 3
- Hcarn:
-
Homocarnosine
- NAA:
-
N-acetylaspartate
- NAAG:
-
N-acetylaspartylglutamate
References
Baslow MH (2000) Functions of N-acetyl-l-aspartate and N-acetyl-l-aspartylglutamate in the vertebrate brain. Role in glial cell-specific signaling. J Neurochem 75:453–459. doi:10.1046/j.1471-4159.2000.0750453.x
Baslow MH (2007) N-acetylaspartate and N-acetylaspartylglutamate. In: Lajtha A (ed) Handbook of neurochemistry and molecular neurobiology, vol 6, 3rd edn. Amino acids and peptides in the nervous system, chap 14, p305–346, pp 418. Springer Science, NewYork. ISBN:978-0-387-30342-0
Baslow MH (2008) The astrocyte surface NAAG receptor and NAAG peptidase signaling complex as a therapeutic target. Drug News Perspect 21(5):251–257
Baslow MH, Guilfoyle DN (2007) Using proton magnetic resonance imaging and spectroscopy to understand brain “activation”. Brain Lang 102(2):153–164. doi:10.1016/j.bandl.2006.06.119
Baslow MH, Suckow R, Sapirstein V, Hungund BL (1999) Expression of aspartoacylase activity in cultured rat macroglial cells is limited to oligodendrocytes. J Mol Neurosci 13(1–2):47–53. doi:10.1385/JMN:13:1-2:47
Baslow MH, Suckow RF, Berg MJ, Marks N, Saito M, Bhakoo KK (2001) Differential expression of carnosine, homocarnosine and N-acetyl-l-histidine hydrolytic activities in cultured rat macroglial cells. J Mol Neurosci 17:87–95. doi:10.1385/JMN:17:3:351
Bowser DN, Khakh BS (2007) Vesicular ATP is the predominant cause of intercellular calcium waves in astrocytes. J Gen Physiol 129(6):485–491. doi:10.1085/jgp.200709780
Burlina AP, Schmitt B, Engelke U, Wevers RA, Burlina AB, Boltshauser E (2006) Hypoacetylaspartia: clinical and biochemical follow-up of a patient. Adv Exp Med Biol 576:283–287. doi:10.1007/0-387-30172-0_20
Coyle JT (1997) The nagging question of the function of N-acetylaspartylglutamate. Neurobiol Dis 4:231–238. doi:10.1006/nbdi.1997.0153
Filosa JA, Bonev AD, Straub SV, Meredith AL, Wilkerson MK, Aldrich RW, Nelson MT (2006) Local potassium signaling couples neuronal activity to vasodilation in the brain. Nat Neurosci 9(11):1397–1403. doi:10.1038/nn1779
Janson CG, McPhee SWJ, Francis J, Shera D, Assadi M, Freese A, Hurh P, Haselgrove J, Wang DJ, Bilaniuk L, Leone P (2006) Natural history of Canavan disease revealed by proton magnetic resonance spectroscopy (1H-MRS) and diffusion-weighted MRI. Neuropediatrics 37:209–221
Kramarenko GG, Markova ED, Ivanova-Smolenskaya IA, Boldyrev AA (2001) Peculiarities of carnosine metabolism in a patient with pronounced homocarnosinemia. Bull Exp Biol Med 132(4):996–999. doi:10.1023/A:1013687832424
Lenney JF, Peppers SC, Kucera CM, Sjaastad O (1983) Homocarnosinosis: lack of serum carnosinase is the defect probably responsible for elevated brain and CSF homocarnosine. Clin Chim Acta 132:157–165. doi:10.1016/0009-8981(83)90243-7
Moffett JR, Namboodiri AMA (2006) Expression of N-acetylaspartate and N-acetylaspartylglutamate in the nervous system. Adv Exp Med Biol 576:7–26. doi:10.1007/0-387-30172-0_2
Perry TL, Kish SJ, Sjaastad O, Gjessing LR, Nesbakken R, Schrader H, Loken A (1979) Homocarnosinosis: increased content of homocarnosine and deficiency of homocarnosinase in brain. J Neurochem 32:1637–1640. doi:10.1111/j.1471-4159.1979.tb02273.x
Schoepp DD, Jane DE, Monn JA (1999) Pharmacological agents acting at subtypes of metabotropic glutamate receptors. Neuropharmacol 38:1431–1476. doi:10.1016/S0028-3908(99)00092-1
Shah AJ, de la Flor R, Atkins A, Stone-Murphy J, Dawson LA (2008) Development and application of a liquid chromatography/tandem mass spectrometric assay for measurement of N-acetylaspartate, N-acetylaspartylglutamate and glutamate in brain slice superfusates and tissue extracts. J Chromatogr B Analyt Technol Biomed Life Sci 876(2):153–158. doi:10.1016/j.jchromb.2008.10.012
Woolley ML, Fricker A-C, Mok MHS, Shah AJ, delaFlor R, Dawson LA, Kew JN (2008) Re-evaluation of N-acetylaspartylglutamate (NAAG) as an agonist at group II mGluRs, and antagonist at NMDA receptors. Neuropharmacology 55(4):630 Abs. # 146
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Baslow, M.H. A novel key–lock mechanism for inactivating amino acid neurotransmitters during transit across extracellular space. Amino Acids 38, 51–55 (2010). https://doi.org/10.1007/s00726-009-0232-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00726-009-0232-0