
Abstract
In contrast to the parent pentadecapeptide gramicidin A (gA), some of its cationic analogs have been shown previously to form large-diameter pores in lipid membranes. These pores are permeable to fluorescent dyes, which allows one to monitor pore formation by using the fluorescence de-quenching assay. According to the previously proposed model, the gA analog with lysine substituted for alanine at position 3, [Lys3]gA, forms pores by a homopentameric assembly of gramicidin double-stranded β-helical dimers. Here, we studied the newly synthesized analogs of [Lys3]gA with single, double and triple substitutions of isoleucines for tryptophans at positions 9, 11, 13, and 15. Replacement of any of the tryptophans of [Lys3]gA with isoleucine resulted in suppression of the pore-forming activity of the peptide, the effect being significantly dependent on the position of tryptophans. In particular, the peptide with a single substitution of tryptophan 13 showed much lower activity than the analogs with single substitutions at positions 9, 11, or 15. Of the peptides with double substitutions, the strongest suppression of the leakage was observed with tryptophans 13 and 15. In the case of triple substitutions, only the peptide retaining tryptophan 11 exhibited noticeable activity. It is concluded that tryptophans 11 and 13 contribute most to pore stabilization in the membrane, whereas tryptophan 9 is not so important for pore formation. Cation–π interactions between the lysine and tryptophan residues of the peptide are suggested to be crucial for the formation of the [Lys3]gA pore.



Similar content being viewed by others
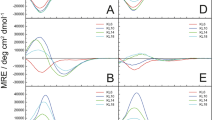
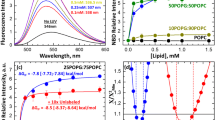

Abbreviations
- gA:
-
Gramicidin A
- [Lys3gA]:
-
gA with lysine in the position 3 instead of alanine
- DPhPC:
-
Diphytanoylphosphatidylcholine
- BLM:
-
Bilayer lipid membrane
References
Aliste MP, MacCallum JL, Tieleman DP (2003) Molecular dynamics simulations of pentapeptides at interfaces: salt bridge and cation-pi interactions. Biochemistry 42:8976–8987
Alvarez C, Mancheno JM, Martinez D, Tejuca M, Pazos F, Lanio ME (2009) Sticholysins, two pore-forming toxins produced by the Caribbean Sea anemone Stichodactyla helianthus: their interaction with membranes. Toxicon 54:1135–1147
Ansorg K, Tafipolsky M, Engels B (2013) Cation-pi interactions: accurate intermolecular potential from symmetry-adapted perturbation theory. J Phys Chem B 117:10093–10102
Antonenko YN, Gluhov GS, Firsov AM, Pogozheva ID, Kovalchuk SI, Pechnikova EV, Kotova EA, Sokolova OS (2015) Gramicidin A disassembles large conductive clusters of its lysine-substituted derivatives in lipid membranes. Phys Chem Chem Phys 17:17461–17470
Arseniev AS, Barsukov IL, Bystrov VF, Lomize AL, Ovchinnikov YuA (1985) 1H-NMR study of gramicidin A transmembrane ion channel. Head-to-head right-handed, single-stranded helices. FEBS Lett 186:168–174
Bai M (2004) Dimerization of G-protein-coupled receptors: roles in signal transduction. Cell Signal 16:175–186
Bamberg E, Apell HJ, Alpes H (1977) Structure of the gramicidin A channel: discrimination between the piL,D and the beta helix by electrical measurements with lipid bilayer membranes. Proc Natl Acad Sci USA 74:2402–2406
Banigan JR, Gayen A, Cho MK, Traaseth NJ (2015) A structured loop modulates coupling between the substrate-binding and dimerization domains in the multidrug resistance transporter EmrE. J Biol Chem 290:805–814
Beechem JM, Brand L (1985) Time-resolved fluorescence of proteins. Annu Rev Biochem 54:43–71
Burkhart BM, Li N, Langs DA, Pangborn WA, Duax WL (1998) The conducting form of gramicidin A is a right-handed double-stranded double helix. Proc Natl Acad Sci USA 95:12950–12955
Busath DD (1993) The use of physical methods in determining gramicidin channel structure and function. Annu Rev Physiol 55:473–501
Chaudhuri A, Chattopadhyay A (2016) Molecular anatomy of an ion channel explored utilizing fluorescence spectroscopy. In: Geddes CD (ed) Reviews in fluorescence 2015. Springer, Heidelberg, pp 353–367
Cosentino K, Ros U, Garcia-Saez AJ (2016) Assembling the puzzle: oligomerization of α-pore forming proteins in membranes. Biochim Biophys Acta 1858:457–466
Das S, Stivison E, Folta-Stogniew E, Oliver D (2008) Reexamination of the role of the amino terminus of SecA in promoting its dimerization and functional state. J Bacteriol 190:7302–7307
Dehez F, Angyan JG, Gutierrez IS, Luque FJ, Schulten K, Chipot C (2007) Modeling induction phenomena in intermolecular interactions with an Ab Initio force field. J Chem Theory Comput 3:1914–1926
Gallivan JP, Dougherty DA (1999) Cation-pi interactions in structural biology. Proc Natl Acad Sci USA 96:9459–9464
Goodchild SC, Angstmann CN, Breit SN, Curmi PM, Brown LJ (2011) Transmembrane extension and oligomerization of the CLIC1 chloride intracellular channel protein upon membrane interaction. Biochemistry 50:10887–10897
Gouaux JE, Braha O, Hobaugh MR, Song L, Cheley S, Shustak C, Bayley H (1994) Subunit stoichiometry of staphylococcal alpha-hemolysin in crystals and on membranes: a heptameric transmembrane pore. Proc Natl Acad Sci USA 91:12828–12831
Hofer N, Aragao D, Caffrey M (2010) Crystallizing transmembrane peptides in lipidic mesophases. Biophys J 99:L23–L25
Johnson RM, Hecht K, Deber CM (2007) Aromatic and cation-pi interactions enhance helix-helix association in a membrane environment. Biochemistry 46:9208–9214
Kamiyama T, Miura T, Takeuchi H (2013) His-Trp cation-pi interaction and its structural role in an alpha-helical dimer of HIV-1 Vpr protein. Biophys Chem 173–174:8–14
Killian JA, Prasad KU, Hains D, Urry DW (1988) The membrane as an environment of minimal interconversion. A circular dichroism study on the solvent dependence of the conformational behavior of gramicidin in diacylphosphatidylcholine model membranes. Biochemistry 27:4848–4855
Koeppe RE, Andersen OS (1996) Engineering the gramicidin channel. Annu Rev Biophys Biomol Struct 25:231–258
Ladokhin AS, Jayasinghe S, White SH (2000) How to measure and analyze tryptophan fluorescence in membranes properly, and why bother? Anal Biochem 285:235–245
Lomize MA, Pogozheva ID, Joo H, Mosberg HI, Lomize AL (2012) OPM database and PPM web server: resources for positioning of proteins in membranes. Nucleic Acids Res 40:D370–D376
Michalek M, Aisenbrey C, Bechinger B (2014) Investigation of membrane penetration depth and interactions of the amino-terminal domain of huntingtin: refined analysis by tryptophan fluorescence measurement. Eur Biophys J 43:347–360
Nicholson LK, Cross TA (1989) Gramicidin cation channel: an experimental determination of the right-handed helix sense and verification of beta-type hydrogen bonding. Biochemistry 28:9379–9385
Peter B, Polyansky AA, Fanucchi S, Dirr HW (2014) A Lys-Trp cation-pi interaction mediates the dimerization and function of the chloride intracellular channel protein 1 transmembrane domain. Biochemistry 53:57–67
Pletneva EV, Laederach AT, Fulton DB, Kostic NM (2001) The role of cation-pi interactions in biomolecular association. Design of peptides favoring interactions between cationic and aromatic amino acid side chains. J Am Chem Soc 123:6232–6245
Rokitskaya TI, Sorochkina AI, Kovalchuk SI, Egorova NS, Kotova EA, Sychev SV, Antonenko YN (2012) The pH-dependent induction of lipid membrane ionic permeability by N-terminally lysine-substituted analogs of gramicidin A. Eur Biophys J 41:129–138
Schroers A, Burkovski A, Wohlrab H, Kraemer R (1998) The phosphate carrier from yeast mitochondria. Dimerization is a prerequisite for function. J Biol Chem 273:14269–14276
Sorochkina AI, Kovalchuk SI, Omarova EO, Sobko AA, Kotova EA, Antonenko YN (2013) Peptide-induced membrane leakage by lysine derivatives of gramicidin A in liposomes, planar bilayers, and erythrocytes. Biochim Biophys Acta 1828:2428–2435
Sorochkina AI, Plotnikov EY, Rokitskaya TI, Kovalchuk SI, Kotova EA, Sychev SV, Zorov DB, Antonenko YN (2012) N-terminally glutamate-substituted analogue of gramicidin A as protonophore and selective mitochondrial uncoupler. PLoS ONE 7:e41919
Strous GJ, Gent J (2002) Dimerization, ubiquitylation and endocytosis go together in growth hormone receptor function. FEBS Lett 529:102–109
Veatch WR, Fossel ET, Blout ER (1974) The conformation of gramicidin A. Biochemistry 13:5249–5256
Vivian JT, Callis PR (2001) Mechanisms of tryptophan fluorescence shifts in proteins. Biophys J 80:2093–2109
Wallace BA (1990) Gramicidin channels and pores. Annu Rev Biophys Biophys Chem 19:127–157
Zhao H, Kinnunen PK (2002) Binding of the antimicrobial peptide temporin L to liposomes assessed by Trp fluorescence. J Biol Chem 277:25170–25177
Acknowledgements
The authors are grateful to Andrei Lomize from the University of Michigan and Tatyana Rokitskaya from Belozersky Institute of Physico-Chemical Biology for fruitful discussions and a help in conducting several experiments. We acknowledge the financial support from the Russian Science Foundation (Grant Number 16-14-10025) to Yuri Antonenko for synthesis of gA derivatives and liposome leakage studies and the financial support from the Division of Biological Infrastructure of the National Science Foundation (Award Number 1458002) to Irina Pogozheva for modeling part of the research.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
All authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Firsov, A.M., Pogozheva, I.D., Kovalchuk, S.I. et al. Effect of Site-Specific Intermolecular Lysine–Tryptophan Interactions on the Aggregation of Gramicidin-Based Peptides Leading to Pore Formation in Lipid Membranes. J Membrane Biol 251, 633–640 (2018). https://doi.org/10.1007/s00232-018-0040-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00232-018-0040-0