
Abstract
Limited experimental evidences are available on the use of peptides as vaccines to boost BCG induced immunity for protection against tuberculosis. The present study therefore evaluated protective efficacy of booster dose of N-terminal peptides of Ag85B, using prime boost approaches in murine model of tuberculosis. Using earlier established subcutaneous murine model of TB in our laboratory, we compared the protective vaccination efficacy of peptides of Ag85B with that of booster dose of whole Ag85B and BCG by evaluating both antibody and cell-mediated immune response. Groups of mice primed by BCG and boosted with Ag85B peptides showed limited pulmonary bacillary burden and reduced lung pathology after challenge with virulent dose of Mycobacterium tuberculosis in mice. Significant levels (p < 0.001) of BCG specific antibodies (anti-BCG, anti-PPD) and T cell-specific cytokines were observed in Ag85B peptides boosted mice compared to Ag85B and BCG. Ag85B and BCG boosted mice however showed significant protection compared to single BCG dose and unvaccinated control groups. Our result suggests that prime boost strategy using N-terminal peptides of Ag85B may improve immunogenicity of BCG against TB. Such peptides may be attractive candidates for boosting waning BCG induced immune response in near future. However study demands further work including improvisation in experimental designs to justify the results.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Tuberculosis (TB) stills remain a global infectious killer, with high rates of mortality and morbidity in developing countries (Luby et al. 2008). From a global perspective, approximately twenty four percent of TB infected population lives in India (Kashyap et al. 2013a, b, c). Only vaccine available for protection against TB in humans is the attenuated strain of Mycobacterium bovis (M. bovis) known as Bacillus Calmette Guerin (BCG) (Senova et al. 2006; Andersen and Doherty 2005).Although BCG provides adequate protection against childhood forms of miliary and meningeal TB, its efficacy for protection against highly infectious forms of adult pulmonary TB remains variable (Agger and Andersen 2002). Natural exposure to environmental mycobacteria is thought to exert an important influence on the immune response, and this may mask or otherwise inhibit the effect of BCG vaccination in developing countries (Brandt et al. 2002; Demangel et al. 2005). In the past decade, there has been a substantial progress in the development of new TB vaccines and many of these have already found their way into clinical trials or pre-clinical development. However, inspite of active research where in over more than 200 vaccine candidates have been evaluated, we have still lacked in developing a molecule that can surpass the safety and Immunogenicity of BCG against TB for use in humans.
BCG protects against childhood manifestation of TB which suggests that it has essential antigenic repertoire for evoking protective immune response but lacks ability to develop enough memory cells for inducing long term protection in adults (Mollenkopf et al. 2004). Current failure of subunit vaccines based on antigens of M. tuberculosis (MTB) to replace/boost BCG induced immunity in adults demands the identification of alternative approaches for effective vaccine development (Tameris et al. 2013). The major task for TB vaccinologists thus lies in developing vaccine candidate that can not only boost BCG induced immunity but also requires less processing by MHC molecules for inducing rapid and efficient immune response. Peptides of major immunogenic antigens of MTB can act as effective vaccine candidate as they can directly bind and interact with MHC molecules to activate T cell response (Allen et al. 1984). Peptide vaccines offer the unique benefit of choosing precisely those epitopes that evoke desired response; simultaneously avoiding motifs that may possibly provoke autoimmune reactions (Chodisetti et al. 2012). Moreover, peptides show least variation in composition, as compared to whole cell-based vaccines where quality control in terms of antigenic composition is difficult. Although peptides are weak immunogens, however using in vitro studies, investigators have earlier demonstrated effectiveness of N-terminal linked peptides of Ag85B for evoking better immune response in BCG vaccinees (Valle et al. 2001; Mustafa et al. 2000). So far, only limited studies are available on the use of MTB Ag85B peptides as vaccine candidates in animal models. In our earlier studies, we have successfully shown effectiveness of heterologous prime boost regimes with Ag85B peptides to boost BCG induced immune response in mice (Husain et al. 2015a, b). However earlier studies investigated prophylactic aspects of peptides based vaccines, in mice, based on simply analyzing increase in BCG induced immunity post boosting. To study protective efficacy of peptide based vaccine, a high dose subcutaneous infection model of TB was further developed in our laboratory (Husain et al. 2015a, b).
Present study was therefore designed to investigate and compare protective efficacy of N-terminal peptides of Ag85B, with that of booster dose of whole Ag85B and BCG using prime boost approaches in murine model of TB.
Materials and Methods
Animal and Human Subject
Female BALB/c mice 6–8 weeks old were purchased from National Institute of Nutrition, Hyderabad and were housed in animal house of Nagpur Veterinary College (Nagpur) under aseptic conditions and provided with food and sterile water. Prior to experimentation, all mice were acclimatized for 15–20 days. For human studies, blood samples were collected from healthy BCG vaccinated participants having no history of pulmonary illness, acid fast bacilli (AFB)/culture negative, Tuberculin skin test (TST)/Quantiferon TB-Gold (QFT-G) negative and seronegative for HIV and used for peripheral blood mononuclear cell (PBMC) isolation.
Vaccine and Reagents
BCG vaccine (Moscow strain) was obtained from Serum Institute of India, Pune and stored at 4–8 °C before use. M. bovis Purified Protein Derivative (PPD) for anti- PPD assay was obtained from Indian Veterinary Research Institute (IVRI), Izatnagar, India. MTB H37Rv antigens ESAT-6 (Rv3875), CFP-10 (Rv3874) were obtained from Colorado State University, USA under the TB Research Materials and Vaccine Testing Contract (NO1-AI-75320). The secondary antibody, rabbit anti-mouse IgG–HRP was obtained from Genei, Bangluru, India.
Designing and Synthesis of Ag85B Peptide
T-cell epitopes of Ag85B specific for CD4 response were identified from reference sequence of Ag85B based on available literature (Valle et al. 2001; Mustafa et al. 2000). Four Peptides P1 (P45–P55) (GLPVEY LQVPSP), P2 (P25–P40) (LPGLVG LAGGAATAGA), P3 (P91–P108) (NG WDINTPAFEW YYQSG), P4 (P110–P126) (IVM PVGGQSSFYS), of Ag85B were designed and synthesized chemically by protocol described elsewhere (Kashyap et al. 2013a, b, c) and evaluated for T-cell activity using PBMC model. Schematic work flow of peptide design and synthesis is given in Fig. 1.

Schematic work flow for peptide designing, synthesis and evlaution
PBMC Isolation and Cytokine Activity
PBMCs from healthy BCG vaccinated donors (n = 15) were isolated as per protocol described by Kashyap et al. (2010). After counting, the cells were cultured in RPMI-1640 medium keeping the concentration at 2 × 105 cells/well and were stimulated with the BCG vaccine (105 CFU/ml) and IL-2 (100 U/well) (Sigma). Cells were then stimulated with peptides (P1–P4) keeping concentration at 25 µg/ml in freund’s incomplete adjuvant (Genei, India) and incubated at 37 °C in an atmosphere of 5 % CO2–95 % air. Cells were taken out at 48 h; supernatant was separated and was analyzed for cytokine activity. The unstimulated cells without BCG/peptide induction were taken as controls. Cells stimulated with BCG only without peptide induction were used as positive control.
Immunization of Animals and MTB Infection
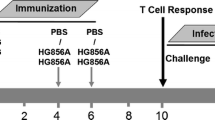
Mice were divided into different experimental sets (n = 10, each set) and primed subcutaneously with 0.1 ml (105 CFU) of BCG vaccine at day 0. A control group of mice (n = 10) were separately maintained and sham immunized with sterile saline. Groups of mice were subsequently boosted with BCG vaccine (102 CFU), Ag85B (25 μg/ml in Freund’s incomplete adjuvant), Ag85B peptides (25 µg/ml of P1/P3 cocktail in Freund’s incomplete adjuvant) in final 500 µl volume twice in the interval of 4 weeks. The MTB H37Rv was grown in 7H9 Middlebrook Broth (Himedia laboratories, India). The bacterial suspension was diluted in phosphate buffered saline (PBS) and adjusted according to the number 1 McFarland scale. 5 weeks after last booster vaccination, all groups of mice were infected by subcutaneous injection with approximately 2 × 106 viable bacilli of MTB H37Rv in a volume of 0.1 ml of 0.9 % sterile sodium chloride solution. 28 days after MTB infection, blood was collected via retro-orbital route from groups of mice and harvested to obtain serum for analysis of antibody response. All the procedures of culturing and infection were done in BSL facilities.
Enumeration of Mycobacteria in Organs
Mice from respective groups (n = 3) were sacrificed 30 and 60 days after MTB infection. Lungs and spleen were isolated, homogenized and serially diluted. These serially diluted homogenates were innoculated in Middle brook 7H9 liquid medium along with oleic acid, albumin, dextrose and catalase (OADC) enrichment and antibiotic supplements in BacT/Alert culture bottles (Biomerieux, France) and incubated at 37 °C in BacT/Alert system (Biomerieux, France) for 30 days. Mycobacterial load in organs was determined in terms of mean time taken by organ cultures from respective groups to become positive in BacT/Alert system as described elsewhere by Kolibab et al. (2014).
Histopathology
A section was prepared from the base of the apical lobe and from the diaphragmatic lobe of the left lung, representing two distinct regions of the organ, ensuring each lobe is well sectioned at the same position for every animal. Duplicate sections were stained with haematoxylin and eosin and Van Gieson in order to aid visualization of fibrous tissue. The two sections were scored in a blinded fashion for the following features: percentage of granulomatous lesion along with necrosis, presence of epithelioid cells and extent of lymphocytic infiltration. A pathological score based on observation of amount infiltration in lung tissues was prepared and interpreted as low (1/+), moderate (++/2) and high (3/+++). Amount of granulomatous lesions were scored in terms of percentage (%) occupied by them in lungs.
Cytokine Estimation
Cytokines (IFN-γ, IL-12, TNF-α) were assessed in organ homogenates and cell supernatants using standard sandwich ELISA method as per manufacturer’s instruction (Bender Med System, Austria). In brief, anti (IFN-γ, IL-12, TNF-α) monoclonal coating antibodies were adsorbed onto microwells. After 2 h of incubation at room temperature, the wells were washed and blocked with 0.5 % BSA in PBS. After 1 h of incubation at room temperature, cell supernatants/spleen homogenates followed by biotin-conjugated anti-cytokine antibodies (IFN-γ, IL-12, TNF-α) were added to the coated wells. After another 2 h of incubation, streptavidin-HRP (horseradish peroxidase) was added to the wells. After 1 h of incubation, streptavidin-HRP was removed by washing and substrate solution reactive with HRP was added to the wells. A coloured product was formed in proportion to the amount of cytokine present in the sample. The reaction was terminated by the addition of 4 N H2SO4 and the absorbance of colour was measured at 450 nm.
Analysis of Antibody Response
Total IgG and antibody titres against BCG and M. bovis PPD in vaccinated and unvaccinated mice samples were evaluated using in house developed ELISA protocol described elsewhere by Husain et al. (2011, 2015a, b). Anti-ESAT-6 and CFP-10 response were evaluated by ELISA protocol described earlier by Kashyap et al. (2013a, b, c).
Statistical Analysis
Data are expressed as mean ± standard deviation (SD). For multiple comparisons, ANOVA was used for obtaining statistical significance. Graphs were plotted using sigma plot version 10.0 and Graph Pad Prism 6 software. P value < 0.05 was considered statistically significant (**) and P < 0.001 for highly significant (***) values.
Results
Cytokine Profile in Human PBMCs
PBMCs from 15 healthy BCG vaccinated participants were isolated and stimulated with BCG vaccine and with different peptides (P1–P4). Figure 2 shows cytokine a) IFN-γ b) IL-12 response in cell supernatants of BCG stimulated PBMCs induced with different peptides. Peptides P1 and P3 were specifically recognized and showed significant T- cell activity (p < 0.001) in cell supernatants compared to peptides P2 and P4, BCG and unstimulated control fraction. Based on the observed T cell response, cocktail of peptides (P1 and P3) were further evaluated for protective efficacy studies in mice model of TB.

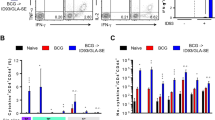
Mean cytokine levels. a IFN-γ, b IL-12 in cell supernatants of short term PBMCs of BCG vaccinees (n = 15) induced with different peptides (P1, P2, P3, P4) of Ag85B. Cells were harvested keeping cell density at 2 × 105 cells/well and stimulated with BCG vaccine. Control represents cells without any induction. PBMCs stimulated with BCG vaccine alone are used as positive control to compare its T-cell activity with that of selected panel MTB Ag85B peptides. Data were analyzed with the Student t test. Individual bars highlight the mean values, and error bars indicate the standard error of the mean; data are representative of 2 independent experiments. (**) represent significant (P1 vs BCG, control, P2, P4) (p < 0.05) and (***) represents highly significant (P3 vs all groups) (p < 0.001) values
Ag85B Peptide Boosted Mice Showed High BCG Specific IgG but Low CFP-10 and ESAT-6 Titres
To determine IgG response, blood was collected 28 days after MTB infection and harvested to obtain serum. Prime boost regimes with Ag85B peptides produced significantly (p < 0.001) higher BCG specific IgG response (Anti-BCG, Anti-PPD) compared to booster doses of BCG and Ag85B post infection (Fig. 3a, b, c). Serum from mice boosted with Ag85B and BCG also showed significantly (p < 0.001) high levels of BCG specific antibodies compared with mice vaccinated with single dose and unvaccinated MTB control. In terms of total IgG response, booster dose of BCG showed higher IgG levels compared to other experimental groups. Antibody titres against CFP-10 & ESAT-6 were also evaluated in different study groups post infection since these antigens are expressed in virulent strains of MTB but absent in all strains of BCG. On similar lines, Ag85B peptide vaccination showed significantly reduced IgG antibodies against ESAT-6 and CFP-10 (p < 0.001) compared to Ag85B, BCG and control groups (Fig. 4a, b). Interestingly booster dose of BCG showed reduced ESAT-6 titre compared to Ag85B, although it was not statistically significant. Infected but unvaccinated mice (MTB control group) showed highest CFP-10 and ESAT -6 antibody titres (p < 0.001), followed by mice vaccinated with BCG alone.

Mean antibody response. a anti-BCG, b anti-PPD and c Total IgG response in serum of vaccinated and unvaccinated mice groups (n = 7, each group). Mice were primed with BCG (105 CFU) at 0th week and boosted with Ag85B peptide (25 μg/ml of P1/P3), Ag85B (25 μg/ml) and BCG (102 CFU) twice in interval of four weeks. Blood was harvested to obtain serum after 28 days post infection to study antibody response. ANOVA was used for multiple comparisons. (**) represent significant (p < 0.05) and (***) represents highly significant (p < 0.001) values

Mean antibody response against ESAT-6 and CFP-10 in serum of vaccinated and unvaccinated control groups (n = 7, each group). Anti-ESAT-6 and CFP-10 titres were estimated 28 days after MTB infection to study correlates of protection, since these antigens are expressed in virulent strain of MTB. ANOVA was used for multiple comparisons Data are shown as ± SD. (**) represent significant (p < 0.05) and (***) represents highly significant (p < 0.001) values
Booster Vaccination with Ag85B Peptides, & Ag85B Showed Reduced CFU and Lung Histopathology Compared to Single BCG Dose and Unvaccinated Controls
30 and 60 days after MTB infection, mice from respective groups (n = 3) were euthanized and organs were isolated and studied for histological changes along with mycobacterial burden in lungs and spleen. Groups of mice boosted with Ag85B peptides showed reduced lung pathology which gradually decreased with progress in infection as compared to booster dose of BCG and Ag85B (Fig. 5a). BCG prime and Ag85B peptide boost combination yielded significantly (p < 0.001) fewer lesions and reduced infiltration in lung post 30 and 60 days after infection (Fig. 5b, c) compared to other groups. Similarly mice boosted with Ag85B showed reduced lung pathology along with fewer lung lesions and infiltration (p < 005) compared to single BCG dose and MTB infection control group. In terms of lesion in lungs, mice boosted with BCG showed fewer lesions at 30 days as compared to other groups but a gradual increase in number of lesions were observed with disease progression. For estimation of bacterial load, 10 fold serially diluted lung and spleen homogenates were inoculated in Middle brook 7H9 liquid medium along with OADC enrichment and antibiotic supplements in BacT/Alert culture bottles (Biomerieux, France) and incubated at 37 °C in BacT/Alert system (Biomerieux, France) for 28 days. Mean time for positivity was taken as correlate for CFU in both organs. Based on the results, we observed that, mice boosted with Ag85B peptides and Ag85B showed significantly (p < 0.05) more time towards MTB culture positivity than single BCG dose and unvaccinated MTB infected control groups, thereby suggesting significantly reduced bacillary load and rapid clearance of MTB in lungs and spleen (Fig. 6a, b). Booster BCG dose also showed reduced CFU compared to unvaccinated control group however its protective efficacy was somewhat less than that observed for Ag85B peptides and Ag85B boosted group.

a Representative Histopathology of lung section of mice stained with hematoxylin and Eosin along with, b extent of infiltration and c lesions. 30 and 60 days after MTB infection, three animals per group were sacrificed and organs were isolated for histopathology. Arrows indicate region of infiltration. Bars indicate mean scores of infiltration and lesions in different mice groups. Pathological score based on observation of amount of infiltration in lung tissues were prepared and interpreted as low (1/+), moderate (++/2) and high (3/+++). Congestion levels were scored based on percentage damage to lung tissue in study groups. (**) represent significant (p < 0.05) and (***) represents highly significant (p < 0.001) values

Shows mean growth time of MTB in lung homogenates of mice (n = 3) collected 30 and 60 days post MTB infection in different study groups. Mice from respective groups were sacrificed and mycobacterial load was determined by inoculating serially diluted lung homogenates in Middle brook 7H9 liquid medium and incubating at 37 °C in BacT/Alert system (Biomerieux, France) for 30 days. Bars represent mean time required for growth in BacT was taken as a correlate of load in respective organs of mice group. Data are shown as ± SD. (**) represent significant (p < 0.05) and (***) represents highly significant (p < 0.001) values
Ag85B Peptide Vaccination Induced Robust Cytokines Response in Organ Homogenates
Figure 7 shows mean cytokines levels of (a) IFN-γ, (b) IL-12, (c) TNF-α in spleen homogenates of different groups of mice at 30 and 60 days post infection. Mice vaccinated with BCG and boosted with Ag85B peptides showed significant (p < 0.001) levels of all cytokines (IFN-γ, IL-12, and TNF-α) after 30 and 60 days post infection compared to other groups. Homogenates from mice boosted with BCG also induced higher IL-12 levels post infection, which was significant (p < 0.05) from Ag85B, and other groups but less than Ag85B peptides. Booster doses of Ag85B and BCG showed higher IFN-γ levels post 30 days after infection but their levels were found to decrease with progress in MTB infection in mice compared to control.

Mean cytokines a IFN-γ, b IL-12 and c TNF-α in spleen homogenates of different groups of mice at 30 and 60 days post MTB infection. Cytokines levels were analyzed in splenocytes stimulated with PPD antigen (25 μg/ml). Data are shown as ±SD. (**) represent significant (p < 0.05) and (***) represents highly significant (p < 0.001) values
Discussion
Inspite of considerable protection against childhood forms of TB, existing BCG vaccination protocol has failed to provide adequate protection against adult pulmonary disease. Superior vaccination strategies that provide long term protection are therefore needed to minimize current TB burden in high endemic regions of developing world (Barreto et al. 2006). Previous studies have shown that repeat vaccination with BCG may be deleterious to protection against TB, and heterologous boost vaccines are likely to be used to enhance specific immunity primed by BCG (McShane and Hill 2005) because BCG is not an effective booster vaccine itself. In the present study we evaluated and compared protective efficacy of N-terminal linked peptides of Ag85B with that of whole Ag85B and BCG using prime boost regimes in murine model of TB. Based on the available literature which suggests efficacy of N-terminal peptides of Ag85B to generate specific CD4 response in BCG vaccines, four peptides spanning different N-terminal regions of Ag85B were designed. Although these peptides were earlier reported to be dominant to induce T- cell response in BCG vaccinees, however, based on heterogeneous population in both studies, these peptides were further screened for CD4 response by evaluating T-cell specific cytokines in PBMCs of BCG vaccinees. PBMCs of BCG vaccinees were stimulated with BCG first and later induced with different peptides of Ag85B. Based on the results we found that peptides P1 and P3 induced significant cytokine (IFN-γ and IL-12) responses in PBMCs of BCG vaccinated individuals compared to unstimulated control thereby suggesting their potential to boost BCG induced immunity. Cocktail of these peptides were further evaluated for protective studies in murine model of TB using prime boost vaccination strategy. Cocktail of peptides were specifically used instead of individual peptides to increase the immunogenicity and booster potential since peptides are reported to be less immunogenic in animals. Mice primed with BCG and subsequently boosted with Ag85B peptides induced good protection rates against virulent dose of MTB bacilli with low bacterial burden and lung pathology and high T-cell specific cytokine responses in spleenocytes which are essential for host defense against TB. Similarly, we observed high BCG specific IgG response in samples of mice boosted with Ag85B peptides. These results thus suggest ability of peptides to efficiently boost BCG induced immunity for protection against TB.
To overcome limitations and improve the efficacy of current TB vaccine, a focus on strategies aiming to boost BCG induced immune response is needed (Martin 2005). The poor performance of BCG, especially in TB endemic areas, is attributed to the interference of environmental mycobacteria in antigen processing and inadequate generation of T-cell memory response (Brandt et al. 2002; Mollenkopf et al. 2004). As mentioned elsewhere, peptides of major immunogenic antigens of MTB can act as effective vaccine candidates as they can directly bind and interact with MHC molecules to activate T cell response. Our study showed that Ag85B peptides were able to subsequently boost and produce enduring immune response in BALB/c mice. Peptide vaccines offer numerous advantages over antigen based vaccines. Peptides are synthetic in nature and do not contain any infectious material. Moreover, peptides show least variation in composition, as compared to whole cell-based vaccines where quality control in terms of antigenic composition is difficult (Purcell et al. 2007). Synthetic peptides have attracted considerable attention as a basis for subunit vaccine design. Peptides play critical roles in determining the magnitude and specificity of cell-mediated (T-cells) and humoral (antibody) immunity, and can contain the minimal sequences necessary for immunomodulation. Delivered correctly, peptides harboring B or T-cell receptor epitopes are sufficient to elicit an effective immune response. Such advantages can be efficiently exploited to design peptide based vaccines in therapeutics. Some of the researchers have studied and showed effectiveness of antigenic peptides as vaccine candidates when used alone or in heterologous prime boost regimes using animal models. Olsen et al. (2000) in their studies showed efficient protection against MTB using 20-mer synthetic construct from ESAT-6 protein. ZUGEL et al. (2001) evaluated immunogenicity of gp-96 associated peptides against intracellular bacteria like MTB and showed that it can act as a potential vaccine candidate. Sugawaraa et al. (2006) have used immunogenic synthetic Ag85A peptide as a booster together with recombinant BCG Tokyo (Ag85A) or Ag85A DNA. They have shown significant improvement in lung pathology and significant reduction in pulmonary CFU. In another studies, investigators have shown that lipidated promiscuous peptide vaccines evoke better immune response and may be alternative to BCG in TB endemic regions (Gowthaman et al. 2012).
An interesting observation from our study was similar protective efficacy of booster doses with BCG and Ag85B to stimulate immune response, which was significantly greater than single BCG dose. Although booster dose Ag85B was able to induce better protection rates compared to BCG based on reduced lung pathology (amount of reduced lesion and infiltration) and organ bacterial load, ability to induce BCG specific IgG response and protective T-cell cytokine response was more or less similar between the two candidates. With current failure of Ag85B based vaccination to boost BCG specific immune response, similar protective efficacy shown by BCG in this context, suggests that subsequent boosting with low dose of BCG may also be used as alternative approach to subunit based vaccine to boost immune response in high TB endemic regions. Although, there is lack of evidences supporting efficacy of booster BCG doses for protection against TB, however in previous studies we have shown immunogenicity of repeat BCG dose to boost immune response using in vitro and animal studies (Kashyap et al. 2010; Husain et al. 2011). Since all attempts to replace BCG have achieved poor success due to lack of knowledge of mechanism of BCG induced immune response, boosting BCG induced immunity by specific T cell epitopes of MTB antigens or BCG remains an ideal approach for future vaccination regimes.
Ideal vaccine should induce number of protective markers. To evaluate protective efficacy of a vaccine, identification of multiple biomarkers which act as surrogate endpoints of protection are needed (Monika et al. 2011). Therefore another aspect of the study was that apart from evaluating T-cell response, we also focused on assessing B cell response. B-cells have been described as an important source of chemokines involved in granuloma development and consequently inhibit mycobacterial dissemination, resulting in recruitment of appropriate cells in the vicinity for the first few weeks after infection (Bosio et al. 2000). During the past few years, there are many published reports which suggest a key role of humoral response in TB immunology. In recent studies it has been shown that antibody responses are essential to contain mycobacterial infection, and there is a synergy and mutual interdependence between cell mediated and humoral immunity (Abebe and Bjune 2009). Zuniga et al. (2012) in their review article have discussed briefly about different cellular and humoral mechanisms involved in control of tuberculosis infection and also suggested a possible role of B cells in modulating immune response in TB. Focus on evaluation of molecules imparting efficient B-cell response apart from T cell immunity is therefore needed as they can be used as effective booster candidates. In our study, we found that peptides of Ag85B induced significant levels of BCG specific IgG response compared with Ag85B and BCG. The high BCG specific IgG response in peptides boosted mice group correlated well with improved protection rates in lungs and reduced bacterial growth. This improved protection rates are suggestive of protective role played by humoral immunity along with already known cell mediated response against MTB infection.
Although our study showed effectiveness of peptide based vaccines to boost BCG induced immunity, the study is also associated with some limitations. The present study evaluated correlates of protection in mice using subcutaneous infection model compared to widely used aerosol model. The main reason was unavailability of aerosol facility at Nagpur Veterinary College. However, to overcome the limitation we had earlier developed and standardized subcutaneous model of TB infection by evaluating multiple markers of TB infection and found that high dose by subcutaneous route induces TB infection specific to aerosol route with characteristic lung pathology (Husain et al. 2015a, b). Another limitation was evaluation of limited number of T-cell specific cytokines which was mainly due to limited sample size. However considering the effectiveness of MTB Ag85B peptides to boost BCG induced immunity, we further plan to investigate by such peptides covering above limitation in future studies for development and screening of improved peptide based vaccines for boosting BCG induced immunity.
To conclude prime boost strategy using N-terminal peptides may improve immunogenicity of BCG against TB. These MTB peptides may be used as attractive candidates to boost waning BCG induced immune response in near future. However study demands further work including improvisation in experimental designs to justify the results.
References
Abebe F, Bjune G (2009) The protective role of antibody responses during Mycobacterium tuberculosis infection. Clin Exp Immunol 157:235–243
Agger E, Andersen P (2002) Novel TB vaccine; towards a strategy based on our understanding of BCG failure. Vaccine 21:7–14
Allen PM, Strydom DJ, Unanue ER (1984) Processing of lysozyme by macrophages: identification of the determinant recognized by two T-cell hybridomas. Proc Natl Acad Sci USA 81:2489–2493
Andersen P, Doherty TM (2005) The success and failure of BCG: implications for a novel tuberculosis vaccine. Nat Rev Microbiol 3:656–662
Barreto M, Pereira S, Ferreira A (2006) BCG vaccine: efficacy and indication for vaccination and revaccination. J Pediatr (Rio J) 82:45–54
Bosio C, Gardner D, Elkins K (2000) Infection of B cell-deficient mice with CDC a clinical isolate of Mycobacterium tuberculosis. J Immunol 164:6417–6425
Brandt L, Feino Cunha J, Weinreich Olsen A et al (2002) Failure of the Mycobacterium bovis BCG vaccine: some species of environmental mycobacteria block multiplication of BCG and induction of protective immunity to tuberculosis. Infect Immun 70:672–678
Chodisetti SB, Rai PK, Gowthaman U et al (2012) Potential T cell epitopes of Mycobacterium tuberculosis that can instigate molecular mimicry against host: implications in autoimmune pathogenesis. BMC Immunol 13:13
Demangel C, Garnier T, Rosenkrands I, Cole ST (2005) Differential effects of prior exposure to environmental mycobacteria on vaccination with Mycobacterium bovis BCG or a recombinant BCG strain expressing RD1 antigens. Infect Immun 73:2190–2196
Gowthaman U, Rai P, Khan N et al (2012) Lipidated promiscuous peptides vaccine for tuberculosis-endemic regions. Trends Mol Med 18:607–614
Husain AA, Kashyap RS, Kalorey DR, Warke SR et al (2011) Effect of repeat dose of BCG vaccine on humoral response in mice model. Indian J Exp Biol 49:7–10
Husain AA, Warke SR, Kalorey DR et al (2015a) Comparative evaluation of booster efficacies of BCG, Ag85B, and Ag85B peptides based vaccines to boost BCG induced immunity in BALB/c mice: a pilot study. Clin Exp Vaccine Res 4:83–87
Husain AA, Daginawala HF, Warke SR et al (2015b) Kashyap RS. Investigation of immune biomarkers using subcutaneous model of M. tuberculosis infection in BALB/c mice: a preliminary report. Immune Netw 15:83–90
Kashyap RS, Husain AA, Morey SH et al (2010) Assessment of immune response to repeat stimulation with BCG vaccine using in vitro PBMC model. J Immune Based Ther Vaccines 8:3
Kashyap RS, Nayak AR, Husain AA et al (2013a) Tuberculosis in India: the continuing challenge. Curr Sci 105:597–606
Kashyap RS, Shekhawat SD, Nayak AR et al (2013b) Diagnosis of tuberculosis infection based on synthetic peptides from Mycobacterium tuberculosis antigen 85 complex. Clin Neurol Neurosurg 115:678–683
Kashyap RS, Nayak AR, Gaikwad S, et al (2013c) Biomarkers in pulmonary and extra pulmonary tuberculosis. In: Proceedings of the international conference on advances in developing affordable invitro molecular diagnostics 73–86
Kolibab K, Yang A, Parra M et al (2014) Time to detection of Mycobacterium tuberculosis using the MGIT 320 system correlates with colony counting in preclinical testing of new vaccines. Clin Vaccine Immunol 21:453–455
Luby SP, Brooks WA, Zaman K et al (2008) Infectious diseases and vaccine sciences: strategic directions. J Health Popul Nutr 26:295–310
Martin C (2005) The dream of a vaccine against tuberculosis; new vaccines improving or replacing BCG. Eur Respir J 26:162–167
McShane H, Hill A (2005) Prime-boost immunisation strategies for tuberculosis. Microbes Infect 7:962–967
Mollenkopf HJ, Kursar M, Kaufmann SH (2004) Immune response to post primary tuberculosis in mice: mycobacterium tuberculosis and Mycobacterium bovis bacille Calmette-Guerin induce equal protection. J Infect Dis 190:588–597
Monika AP, Sergiusz M, Elzbieta K et al (2011) Proteomics for development of vaccine. Proteomics 74:2596
Mustafa AS, Shaban FA, Abal AT et al (2000) Identification and HLA restriction of naturally derived Th1-cell epitopes from the secreted Mycobacterium tuberculosis antigen 85B recognized by antigen-specific human CD4(+) T-cell lines. Infect Immun 68:3933–3940
Olsen AW, Hansen PR, Holm A et al (2000) Efficient protection against Mycobacterium tuberculosis by vaccination with a single subdominant epitope from the ESAT-6 antigen. Eur J Immunol 30:1724–1732
Purcell AW, McCluskey J, Rossjohn J (2007) More than one reason to rethink the use of peptides in vaccine design. Nat Rev Drug Discov 6:404–414
Senova L, Harbacheuski R, Moreira AL et al (2006) Evaluation of the Mtb72F polyprotein vaccine in a rabbit model of tuberculous meningitis. Infect Immun 74:2392–2401
Sugawaraa I, Udagawaa T, Taniyamab T (2006) Protective efficacy of recombinant (Ag85A) BCG Tokyo with Ag85A peptide boosting against Mycobacterium tuberculosis-infected guinea pigs in comparison with that of DNA vaccine encoding Ag85A. J tube 87:94–101
Tameris MD, Hatherill M, Landry BS et al (2013) Safety and efficacy of MVA85A, a new tuberculosis vaccine, in infants previously vaccinated with BCG: a randomised, placebo-controlled phase 2b trial. Lancet 381:1021–1028
Valle MT, Megiovanni AM, Merlo A et al (2001) Epitope focus, clonal composition and Th1 phenotype of the human CD4 response to the secretory mycobacterial antigen Ag85. Clin Exp Immunol 123:226–232
Zugel U, Sponaas A, Neckermann J et al (2001) gp96-Peptide vaccination of mice against intracellular bacteria. Infect Immun 69:4164–4167
Zuñiga J, Torres G, Santos M et al (2012) Cellular and humoral mechanisms involved in the control of tuberculosis. Clin Dev Immunol 2012:193923
Acknowledgments
AAA would like to thank Indian Council of Medical Research (ICMR), Govt of India, for Award of Senior Research Fellowship. Authors acknowledge Ms. Prachi Bapat, for her assistance in statistical analysis.
Funding
This study was funded by Central India Institute of Medical Sciences as part of its In house study.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
All Authors declare that they have no conflict of interest
Ethical Approval
All protocols of animal experiments were approved by Animal Ethics Committee of Central India Institute of Medical Sciences (CIIMS), Nagpur. All applicable International, National, and/or Institutional guidelines for the care and use of animals were followed. All procedures performed in studies involving human participants were in accordance with the ethical standards of the Institutional Ethics committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Informed Consent
Written consent were obtained from each participant and oral explanation about the research work was given to them.
Additional information
Girdhar M. Taori—Deceased
Rights and permissions
About this article

Cite this article
Husain, A.A., Daginawala, H.F., Warke, S.R. et al. Heterologous Prime Boost Regimes with N-terminal Peptides of Ag85B Induces Better Protection than Ag85B and BCG in Murine Model of Tuberculosis. Int J Pept Res Ther 22, 143–153 (2016). https://doi.org/10.1007/s10989-015-9490-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10989-015-9490-8