
Abstract
Laguna del Sauce, one of the main drinking water sources in Uruguay, is an eutrophic shallow lake with high temporal variation of inorganic turbidity caused by extreme wind events. During low turbidity periods, high phytoplankton biomass can be reached, frequently associated to cyanobacteria blooms, which can cause interferences in the water supply. In this study, we assessed the environmental drivers of cyanobacteria dominance, composition, and nitrogen-fixation behavior. For this, we analyzed the spatial and temporal phytoplankton composition, physical and chemical variables performing weekly samplings during two summers: 2015–2016 and 2016–2017. When inorganic turbidity was high (above 30 NTU), phytoplankton biomass was controlled, below this threshold, temperature, secchi depth and nutrients played key factors controlling cyanobacteria biomass and composition. Blooms of N2-fixing cyanobacteria (Dolichospermum crassum, Aphanizomenon gracile, and Cuspidothrix issatschenkoi) were promoted by low N:P ratios (average 11.5) and wide TN range (286–1300 µg l−1). Non-heterocystous cyanobacteria blooms occurred above TN 1000–1200 µg l−1. The N2-fixing behavior (heterocytes:vegetative cell ratio) depended on TN, it was highest at low TN (< 700 µg l−1) but null above ca. 1000 µg l−1. While low inorganic turbidity allowed cyanobacteria blooms in Laguna del Sauce, its composition and N2-fixation behavior depended on the TN and TP levels.
Similar content being viewed by others


Introduction
The temporal and spatial dynamics of phytoplankton are controlled by several external factors, including resource availability, grazing, sinking, and resuspension, among others (Margalef, 1978; Reynolds, 2006). Likewise, different processes of the internal successional dynamics of phytoplankton can also affect the light and nutrient availability. In simple terms, the spatial and temporal patterns of the phytoplankton community in a shallow lake are the result of the interaction between external factors (allogenic) and the internal dynamics of the phytoplankton community (autogenic) (Reynolds, 2006).
The adverse consequences of eutrophication, particularly the occurrence of harmful cyanobacteria blooms, and their negative effects in the provision of key ecosystem services for the human well-being demand a higher capacity to predict and mitigate their occurrence (Paerl et al., 2016). In the last decades, multiple approaches have arisen, which try to understand the temporal phytoplankton dynamics and predict the species assemblage, combining different morphological and functional approaches and attributes (Reynolds et al., 2002; Weithoff, 2003; Kruk et al., 2010). Some contributions have focused in the cyanobacteria component in terms of their ecotypes (e.g., scum forming or dispersive blooms) (Chorus & Bartram, 1999).
The shallow lake analyzed in the present research, Laguna del Sauce, presents contrasting spatial ecosystem configurations, including regions dominated with dense submerged macrophytes and regions with open water free of macrophytes. At the same time, Laguna del Sauce constitutes the second drinking water supply in Uruguay (in terms of the number of people it supplies, 160.000 inhabitants) (INE, 2011).
In Laguna del Sauce, an important part of the phytoplankton temporal biomass dynamics, and its morpho-functional composition (MBFG sensu Kruk et al., 2010), is controlled by a marked variation in the water turbidity and consequently of light availability. These fluctuations respond to extreme wind and/or precipitation events (Crisci et al., 2017a, b). Abrupt shift towards high turbidity occurs in a period of hours and persists for periods over a year and a half, independent of seasonality (Crisci et al., 2017a). In other words, during periods of high inorganic turbidity, phytoplankton development is low (including cyanobacteria), probably because of the strong light constraint. However, when turbidity is low, a high oscillation in the phytoplankton community in general, and of cyanobacteria in particular, occurs. Indeed, during the low turbidity period of 2015, Laguna del Sauce underwent a major cyanobacteria bloom event of the heterocystous nitrogen fixing (heterocystous N2-fixing) Cylindrospermopsis raciborskii (Seenayya & Subba, 1972). The latter caused important water supply crisis, mostly because of taste and odor problems in the drinking water, which caused a huge economic loss and societal turmoil for the country. This bloom was toxic too (saxitoxin), yet the toxin was controlled at the drinking plant facility using activated carbon.
Remarkably, in this ecosystem, during low turbidity periods, when cyanobacteria bloom, its composition is recurrently dominated by heterocystous N2-fixing species (Order Nostocales) and less frequently by non-heterocsytous N2-fixing cyanobacteria (e.g., Microcystis complex) (González-Madina et al., 2017). This poses the question if the success of heterocystous N2-fixing cyanobacteria in this ecosystem relies on its ability to fix atmospheric nitrogen. It is widely recognized that blooms of heterocystous N2-fixers recurrently occur behaving as N2-fixers (e.g., Ferber et al., 2004; Kumar et al., 2010), yet blooms can also occur behaving as non-N2-fixers due to lack of heterocyte development in the filaments (Jacobsen & Simonsen, 1993; Wolk et al., 1994; Zhang et al., 2006). Indeed, information about the N2-fixing behavior in Nostocales can be inferred from the density of heterocytes (cell where fixation occurs, which differentiates from vegetative cells, usually under N scarcity) (de Tezanos Pinto & Litchman, 2010) or the ratio between heterocytes and vegetative cells (Yema et al., 2016), among other methods.
The drinking water crisis supply that occurred in 2015 promoted the implementation of new monitoring programs in order to understand the ecosystem dynamics and to develop an early warning system for the management of the drinking water supply (Crisci et al., 2017b; González-Madina et al., 2017). In this study we analyzed, during two summer cycles (2015–2016 and 2016–2017), the phytoplankton dynamics on a weekly basis, both in the vegetated and in the open water regions of Laguna del Sauce. We aimed at addressing the following questions:
-
Do the resource constraints (light and nutrients) and the temperature variability explain the temporal cyanobacteria dynamics, composition and N2-fixing behavior?
-
Why is the phytoplankton community recurrently dominated by heterocystous N2-fixing cyanobacteria species?
-
Under which environmental conditions is the N2-fixing behavior active? Do all species of N2-fixers respond in a similar way to the environmental conditions?
-
Are cyanobacteria from the order Nostocales dominant only because they fix N2 or other morpho-functional (e.g., cell or filament morphology) and ecophysiological attributes (e.g., growth rate, light traits, dormant cells) provide fitness to this group in this environment?
-
Does the N2-fixation determine autogenic succession mechanisms or is the phytoplankton dynamic mostly governed by external drivers?
Materials and methods
Study area
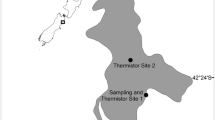
Laguna del Sauce (34°43′S, 55°13′W) is located in the Department of Maldonado, Uruguay (Fig. 1) and provides the ecosystem services of drinking and recreational use of water. This shallow system was a coastal lagoon until 1947, when a dam was built at its mouth with the purpose of maintaining a stable water level for the Air Force Base constructed after World War II by the USA Army.

Laguna del Sauce system, with its three sub-systems: Laguna del Sauce without submerged vegetation, Laguna de los Cisnes with partial submerged plant cover and Laguna del Potrero with more than 60% of its bottom covered with submerged plants. Black dots indicate the six sampling sites
At present, Laguna del Sauce is composed by three connected shallow subsystems: Laguna del Sauce (4045 ha), Laguna de los Cisnes (205 ha), and Laguna del Potrero (411 ha) (Fig. 1). Laguna del Sauce is an open water system without submerged vegetation, most of the area of Laguna del Cisne is free of submerged vegetation while Laguna del Potrero is very shallow and dominated by submerged vegetation—mainly Egeria densa (Planch, 1849) and Ceratophyllum demersum (Linneo 1753) (Mazzeo et al., 2010)—covering more than 60% of the bottom surface, at least for the last 10 years. The main tributaries are the Pan de Azúcar and the Sauce streams, and the ecosystem drains into the Atlantic Ocean through the Potrero stream (Inda & Steffen, 2010) (Fig. 1).
The mean water input is 285.4 hm3 (cubic hectometers) per year, and the water output is 88 hm3, out of which 18 hm3 corresponds to the water withdrawal posed by the drinking water service administrator (OSE-UGD) (Crisci et al., 2017a). Laguna del Sauce is currently the second source of drinking water in Uruguay (it supplies approximately 160,000 permanent inhabitants; this figure more than doubles during the summer season when high-income tourists visit Punta del Este beaches) (INE, 2011).
This ecosystem is eutrophic (average TP: 80 µg l−1 and TN: 200–1000 µg l−1), with different responses depending on the subsystem considered (Inda & Steffen, 2010; Rodríguez et al., 2010). In Laguna del Sauce, complex cyanobacteria blooms have been observed since the 1960s, probably related to enhanced nutrient inputs due to agricultural activity, cattle production intensification, growth of urbanization and tourism (Mazzeo et al., 2010; Pacheco et al., 2010; Rodríguez et al., 2010) and low turbidity (Crisci et al., 2017a). Recently, cyanobacteria blooms intensified (higher frequency and higher biomass, wider distribution, increased persistence, and toxic events).
Sampling and analysis of physical, chemical, and biological variables
Within the framework of the early detection cyanobacteria monitoring program in Laguna del Sauce—carried out by the CURE-Faculty of Sciences (UdelaR), SARAS Institute and OSE-UGD (drinking water supply company)—weekly samplings were carried out during two summer seasons (2015–2016 and 2016–2017). Six sites were sampled in Laguna del Sauce complex encompassing the spatial heterogeneity of the system (Fig. 1). In each sampling station, light penetration was measured with the Secchi disk (SD). The following environmental variables were measured in situ: temperature, conductivity, pH, turbidity, dissolved oxygen, chlorophyll a, and phycocyanin, using a YSI 6600 multiparameter probe. Simultaneously, three independent samples were collected, using a sampling of 10-cm-diameter tube, by vertically integrating the water column. Each sample was considered as a replicate. A fraction of these samples, kept cold and in darkness, was used for the physical and chemical analyses in the laboratory, including total suspended solids by drying at 105°C for 12 h, organic matter (OM) content by weight difference at 550°C, ammonium (NH4, Muller & Wiedemann, 1955), nitrate (NO3), soluble reactive phosphorus (SRP, Murphy & Rilley, 1962), total nitrogen and phosphorus (TN, TP, Valderrama, 1981), and chlorophyll a by extraction in cold ethanol and spectrophotometry (Nusch, 1980).
The phytoplankton replicates were fixed in acidified Lugol and were quantified in the laboratory in sedimentation chambers of 10 or 20 ml, using an inverted microscope at ×400 and ×1000 (Utermöhl, 1958; Guillard, 1978). Random fields were used (Uhelinger, 1964; Utermöhl, 1985) and at least 100 organisms of the most frequent species were counted (Lund et al., 1958). Samples for phytoplankton qualitative analyses (identification and morphological measurement of organisms) were taken with a phytoplankton net (20 μm mesh) and fixed in acidified formaldehyde (formol 40% + acetic acid). From these samples, phytoplankton organisms, colonies, or cells larger than 2 μm were identified and measured, under light microscope (Nikon Eclipse 50 i) at ×400 and ×1000 magnification. For the mentioned measurements, a CMOS 5.0 Mpx camera and the software Micrometrics® Premium Edition, previously calibrated for the different magnifications, were used. The individual volume of the organisms was calculated considering to their morphological traits (length, width) and geometric form according to Hillebrand et al. (1999), measuring at least 30 organisms of each species. The biovolume (biomass) of each species was calculated as its density multiplied by its volume.
To assess the N-fixing behavior of the Nostocales species (species that fix N in heterocytes), samples with biovolume ≥ 1 mm3 l−1 were analyzed, at contrasting TN concentrations. For each sample and species, the heterocyte and vegetative cell numbers per filament were counted in 30 filaments randomly selected. For species in which the cross walls were impossible to differentiate, the filament length was measured, and its vegetative cell length was estimated from updated literature (Komárek, 2013); next the number of vegetative cells per filament was computed. The average heterocyte/vegetative cell ratio per filament was calculated as a proxy for N2-fixing behavior: the higher the ratio the higher the investment in N-fixation. For vegetative cell and heterocyte counts, a direct Olympus CX31 was used; photographs were taken at ×40 and an Infinity 1 camera (infinity analyze software) was used.
Data analysis
In order to analyze which environmental variables explained the phytoplankton composition, first a redundancy analysis (RDA) was performed, to analyze the variability in data gradients. Next, a canonical correspondence analysis (CCA) was performed, considering the physical and chemical variables as explanatory variables, and the biomass of the species as a response variable. The significance of the CCA model was tested by Monte Carlo with 999 unrestricted permutations. The significance of the canonical axes of ordering and of each variable included in the model was also tested, assuming a maximum value of 0.05 in each case.
Patterns of biomass for the different cyanobacteria functional groups (total cyanobacteria, heterocystous N2-fixers, and non-heterocystous N2-fixers) in relation to TN and TP were examined using graphical representation (heat maps graphs). The ratio between TN and TP was calculated using a standard major axis (SMA) analysis. Cross-correlations between heterocystous N2-fixers and total nitrogen were performed to analyze possible cause-effect correspondence between these variables. All statistical analyses were performed in R (R-Core Team 2016) with the packages “vegan” for multivariate analysis, ggplot2 and lmodel2 for the graphical representation and linear regression, respectively.
Results
The two summer periods monitored (2015–2016 and 2016–2017) were characterized by very different phytoplankton biomass (biovolume) and composition (Fig. 2). During the first summer, we observed important temporal variations in the phytoplankton biomass in Laguna del Sauce (sites Sauce Norte—SN, Sauce Sur—SS, and Bahía Mallorquina—BM) (Fig. 2A). Biomass in these sites always surpassed the alert level 3 (2 mm3 l−1) and were markedly higher than the biomass in Laguna del Cisne and Potrero (Fig. 2A). Nevertheless, towards the end of the summer (from 10/03/2016 onwards), the phytoplankton biomass decreased markedly throughout the whole system (Fig. 2A).

Spatial and temporal patterns of phytoplankton biomass (biovolume) across the six sites sampled in Laguna del Sauce complex, for the two summers studied A December 2015 to April 2016 and B December 2016 to April 2017
During the second summer, the spatial pattern was completely different, with higher phytoplankton biomass in Laguna del Potrero (dominated by submerged macrophytes) followed by Laguna del Cisne, mostly during the first half of the summer (Fig. 2B). For the remaining part of the summer, the whole lake showed very low biomass (Fig. 2B).
Phytoplankton composition also varied markedly between both summers. Cyanobacteria biomass was three orders of magnitude higher in the first than in the second summer (Fig. 3A, B). In the first summer, during the 10 weeks with cyanobacteria prevalence in Laguna del Sauce sub-system (SN, SS and BM), in six occasions we found a major contribution or total dominance of N2-fixing cyanobacteria (30–100%) (Fig. 3C). The heterocyte to vegetative cell ratio was very low (proxy of low fixation investment) during the first 5 weeks (0.006 ± 0.02), but about three times higher in weeks 6 and 7 (0.018 ± 0.001). The heterocystous N2-fixing species which bloomed with higher magnitude (biomass ≥ 1 mm3 l−1) were Dolichospermum crassum, Cuspidothrix issatschenkoi (Rajaniemi et al., 2005), and Aphanizomenon gracile (Lemmermann, 1907). The latter two species coexisted in the phytoplankton assemblage during the first three weeks, whereas Dolichospermum crassum bloomed afterwards and in combination with non-N2-fixing colonial cyanobacteria from the Microcystis complex (Microcystis aeruginosa (Kützing, 1846), M. panniformis (Komárek et al., 2002), and Sphaerocavum brasiliense (Azevedo et al., 2003), but not with other N-fixers. During the second summer (2016–2017), cyanobacteria biomass was very low (Fig. 3B), with alternation of heterocystous N2-fixing and non-N2-fixing cyanobacteria (Fig. 3D). The higher cyanobacteria biomass during this period was observed in Laguna del Potrero (vegetated dominated system) reaching the alert level 3 (28.66 mm3 l−1) not in Laguna del Sauce.

Total (A, B) and relative (C, D) Cyanobacteria biomass in open water system of Laguna del Sauce (sites Sauce Norte, Sauce Sur and Bahía Mallorquina) in each of the dates sampled across the two summer periods, December 2015–April 2016, and December 2016–April 2017. In subsets A and B, the points represent the average biomass value (including all replicates). The error bars denote standard deviation among sites monitored in the open water system and replicates analyzed. Note the different scales in subsets A and B
The first summer showed lower turbidity compared to the second summer (Table 1). Towards the end of the first summer (from 2/3/2016 onwards), we observed a sharp increase in the turbidity; from ca. 5–20 NTU to 25–35 NTU. After this, the phytoplankton total biomass markedly dropped (Fig. 2A). Laguna del Potrero showed the highest Secchi disc and lowest turbidity values compared to the other systems (Table 1).
TN was higher during the first summer than in the second, whereas TP remained rather similar across summers (Table 1). Contrary, nitrate and SRP were lower and ammonium higher during the first summer, than in the second summer (Table 1). At spatial scale, the lowest nutrient concentrations were observed at Laguna del Potrero (Table 1).
The canonic correspondence analysis (CCA), encompassing both summer cycles, indicated that the principal environmental factors which explained the temporal variations in phytoplankton composition of Laguna del Sauce sub-system (sites SN, SN and BM) were turbidity, Secchi disc depth, SRP concentrations, and temperature (Fig. 4). Turbidity was inversely linked to the secchi disc depth and temperature, and directly linked with SRP (Fig. 4). All cyanobacteria (heterocystous N2-fixing and non-N2-fixing) were associated with periods of higher temperature, higher Secchi depth but lower turbidity and SRP (Fig. 4). Most heterocystous N2-fixers (Cuspidothrix issatschenkoi, Aphanizomenon, and Raphidiopsis mediterranea) were associated to higher values of Secchi disk depth (Fig. 4). These species bloomed jointly during the onset of the summer of 2015–2016. The heterocystous N2-fixing cyanobacteria Dolichospermum crassum, conversely, were found in similar scenarios to those of non-N2-fixing species of the Microcystis complex (Microcystis aeruginosa, M. panniformis, and Sphaerocavum brasiliense), linked to lower Secchi disk depth (Fig. 4). Dolichospermum crassum bloomed separated in time from the other heterocystous N2-fixers. Dinoflagellates and cryptophytes biomass were linked to high turbidity and high SRP concentrations but low temperature and Secchi disc depth (Fig. 4).

CCA biplot pooling data on both summer periods (2015–2016 and 2016–2017) from the open water system (sites SN, SS and BM). The first two axes of the ordination explained the 76.8% of the variance of the compositional data (λ1 = 42.0%; λ2 = 34.8%). The model was significant (F = 1.24; P = 0.048) as well as the axis of the ordination (λ1: F = 2.04, P = 0.02; λ2: F = 1.70, P = 0.05) and the indicated variables (P < 0.05)
When analyzing the summer periods separately (Fig. 5), in the first summer, in addition to the environmental variables mentioned before, the fluctuations associated with TN and total suspended solids (TSS) were significantly associated with the temporal variations observed in the cyanobacteria assemblage (Fig. 5A). The cyanobacteria able to form scums on the surface (Dolichospermum crassum and species within the Microcystis complex) coincided with the highest values of TSS in the water column and lower transparency (Fig. 5A). The links of cyanobacteria composition and abundance fluctuations with TN are analyzed in detail below.

CCA biplot of each summer: A 2015–2016 and B 2016–2017. In the biplot A the two first axis of the ordination explained 66.7% of the data variance (λ1 = 37.5%; λ2 = 29.2%). The model was significant (F = 1.89; P = 0.02), as well as the axis of the ordination (λ1: F = 4.26, P = 0.002; λ2: F = 3.31, P = 0.01) and the variables considered (P < 0.05). In the biplot B the two first axis of the ordination explained the 67.3% of the data variance (λ1 = 42.3%; λ2 = 25.1%). The model was significant (F = 2.3401, P = 0.001), as well as the ordination axis (λ1: F = 3.9618, P = 0.001; λ2: F = 2.3480, P = 0.003) and the variables considered (P < 0.05). All the data belong to the open water system (sites SN, SS and B)
During the second summer, although cyanobacteria relative biomass was very low, we also found a positive link between cyanobacteria and increased temperature (Fig. 5B). Nutrient concentrations (TN, N–NH4+, and SRP) were negatively linked to temperature but positively linked to cryptophyte occurrence (Fig. 5B). N–NH4+ and TN lacked a straightforward relation; higher values of N–NH4+ coincided with the presence of green algae (Monoraphidium spp.) and euglenoids (Euglena). In this summer, turbidity lacked explanatory power for the temporal variations in phytoplankton composition of Laguna del Sauce (Fig. 5C).
The highest total cyanobacteria biomass in Laguna del Sauce sub-system (sites SS, SN, and BM) occurred at high TN (> 800 µg l−1) and TP between 50 and 100 µg l−1 (Fig. 6A). The different cyanobacteria functional groups showed contrasting distributions along the TN gradient. That is, heterocystous N2-fixing cyanobacteria achieved its highest biomass at intermediate TN concentrations (ca 800 µg l−1) (Fig. 6B), whereas the non-N2-fixing cyanobacteria achieved its highest biomass only at high TN (> 1000 µg l−1) (Fig. 6C).

Cyanobacterial biomass (mm3 l−1) distribution along the TN and TP gradients in both summers, in the open water system (sites SN, SS, and B). A Total cyanobacteria (heterocystous and non-N2-fixing), B heterocystous N2-fixing cyanobacteria and C non-N2-fixing cyanobacteria
Figure 7 shows the biomass distribution and N2-fixing behavior of the dominant heterocystous N2-fixing cyanobacteria species along the TN and TP gradients during the first summer (2015–2016 which was the summer dominated by cyanobacteria), in all six sites analyzed. These species occurred only above the thresholds of 500 µg l−1 TN and 50 µg l−1 TP (Fig. 7). The N2-fixing behavior was plastic and strongly regulated by the TN concentration: the highest average ratio of heterocyte to vegetative cell per filament (proxy of N fixation, where the highest values point to higher N2-fixation) occurred at lower TN concentrations (below 1000 µg l−1 TN) and it was zero above 1000 µg l−1 TN (Fig. 7). The threshold of the onset of N2-fixation varied among the heterocystous N2-fixing species: in Aphanizomenon and Dolichospermum it initiated at TN < 1000 µg l−1, while in Cuspidothrix at TN < 750 µg l−1 (Fig. 7). In general, the biomass of the heterocystous N2-fixers was lower when behaving as N2-fixers than when behaving as non-N2-fixers, except in Dolichospermum (Fig. 7). The regression (type II, SMA) between the TN and TP concentrations showed a slope of 11.5. The highest biomass was found above the estimated slope, at low relative TP to TN concentrations, where P limitation would be expected. In this sense, the CCA result confirmed that higher cyanobacteria biomass was associated to low SRP concentrations (Fig. 4).

Biomass distribution and N2-fixing behavior (ratio of heterocytes to vegetative cells) of the dominant heterocystous N2-fixing species, along the TN and TP gradients, for all sites analyzed (AS, BM, SN, SS, C, P), in the summer dominated by cyanobacteria (2015–2016). Note that the higher the ratio of heterocytes to vegetative cells, the higher the amount of heterocytes (proxy of higher fixation); when the ratio is zero there is absence of heterocytes and hence lack of N2-fixation
Finally, the biomass of the heterocystous N2-fixers (regardless of their fixing behavior) and the TN concentration were coupled most of the time, as reveled by the auto-correlation analysis for the summer of 2015–2016 (Fig. 8).

Cross-correlation analysis of the temporal series corresponding to the biomass of the N2-fixers (black line) and the TN content (red line). The green line denotes the 95% confidence interval. The x axis corresponds to the number of days since the onset of the sampling period (from 31/12/2015 to 24/2/2016) in the summer when cyanobacteria dominated the phytoplankton
Discussion
The results obtained in the current study allow us to address the questions posed in the introduction. Cyanobacteria reached high biomass when the turbidity was low and the temperature was high. Besides these external controls, nutrients, in particular total nitrogen and also phosphate played a significant role in explaining phytoplankton variation in this ecosystem, as found by Crisci et al. (2017a, b). The emerging pattern was that the highest TN concentrations coincided with the highest biomass of cyanobacteria, both of heterocystous N2-fixers and non-N2 fixers, underscoring the risks caused by high nutrient availability. This pattern is very similar to the one observed by Dolman et al. (2012) in 100 lakes in Germany, in terms of biomass. The TP concentrations observed corresponded to eutrophic and hypereutrophic systems (OECD, 1982), yet increases in biomass seemed more affected by TN and the N:P ratio, than to TP alone. As found by Moisander et al. (2012) we found that different cyanobacteria communities gain benefit under distinct N/P scenarios during the cyanobacteria bloom period. Our study, in addition to confirm the findings of Dolman et al. (2012) in terms of biomass, expands the understanding of cyanobacteria ecology as it shows how its physiology (N2-fixing behavior) changes along nutrient gradients (see below).
The maximum cyanobacteria biomass (e.g., 20 mm3 l−1) occurred in the non-N2-fixing colonial cyanobacteria of the complex Microcystis. Though heterocystous N2-fixing cyanobacteria reached lower maximal biomass (12 mm3 l−1), most of the blooms were composed by this functional group. The N2-fixing behavior was plastic: at TN concentrations higher than 1000 µg l−1 all the species behaved as non-N2-fixers (lack of heterocyte presence), whereas at values of 600 µg l−1 TN all species were investing strongly in fixation. Finally, at concentrations of about 800–900 µg l−1 TN, heterocytes were present with low frequencies. The field evidence suggested that fixation is a continuous process between the extremes of lack of fixation and maximum fixation, where fixation complemented the nitrogen needs. Studies assessing the number of heterocytes also show that heterocystous N2-fixers may or may not invest in fixation during blooms of high magnitude (e.g., Jacobsen & Simonsen, 1993; Padisák & Istvánovics, 1997). Other environmental variables, that affect N2-fixation include a higher demand in macro (P) (Vitousek et al., 2002) and of light (Mugidde et al., 2003; Agawin et al., 2007; de Tezanos Pinto & Litchman, 2010), were in sufficient supply in Laguna del Sauce during blooms presenting heterocytes. It is worth highlighting that Laguna del Sauce presented medium to high TP concentrations that the SRP showed an inverse relation with cyanobacteria biomass and that SRP was always above values considered limiting for phytoplankton growth (Reynolds, 2006).
The dominant heterocystous N2-fixing cyanobacteria found—Dolichospermum crassum, Aphanizomenon gracile, and Cuspidothrix issatschenkoi—span 3 of the 10 planktonic genera within Nostocales (Komárek, 2013) which are potentially capable of producing a wide range of cyanotoxins (Carmichael & Boyer, 2016). The laboratory studies of Yema et al. (2016) found that Dolichospermum can reach high biomass both in lack of supplied N and at high N supply, yet express heterocytes only during N-limitation (Yema et al., 2016). Our field results support the results observed in the laboratory, both in terms of biomass and heterocyte presence, which only occurred at TN < 1000 µg l−1. Aphanizomenon gracile and Cuspidothrix issatschenkoi behaved similar to Dolichospermum in terms of heterocyte presence along TN gradients. This result suggests that the heterocyte-to-vegetative cell ratio can be an adequate proxy of N-fixation for these species.
Aphanizomenon and Cuspidothrix, compared to Dolichospermum, only reached high biomasses at high TN concentrations, when they were behaving as non-N2-fixers (lack of heterocyte presence). Our field evidence demonstrates that, from a functional point of view, a same functional group can behave either as fixer and non-fixer, and that the fitness provided by N2-fixation varies across species. Indeed Moisander et al. (2012) found that Anabaena sp. improve its fitness under diazotrophic conditions while C. raciborskii has high fitness under both diazotrophic and non-diazotrophic conditions. These findings may explain the different biomass distributions of several heterocystous N2-fixing species observed in Dolman et al. (2012). Regardless of the TN concentration, other traits different from N-fixation can provide fitness to the group such as light traits (see below). Cylindrospermopsis raciborskii, though it was the species responsible for the massive bloom and crisis in autumn 2015, during the current study, it occurred rarely and with very low biomasses (less than alert level 1).
The frequent blooms of heterocystous N2-fixing cyanobacteria in Laguna del Sauce probably respond to the low average TN concentration (mean, 700 µg l−1, SD, 200 µg l−1) and low N:TP ratio (11.5). As found by Pádisak (1992) for a lake in Hungary, increased P availability leads to a relative N-deficiency, which can be supplied by N fixation.
The cross-correlation analysis between the biomass of heterocystous N2-fixing cyanobacteria and TN evidences (a) an important internal variability and (b) a coupling of both variables. These results, in addition to those in the multivariate analysis, seem to indicate that most of the variability of TN is governed by external factors and to a lesser extent by the N2-fixing behavior of the heterocystous N2-fixing cyanobacteria. In only two of the 14 samplings, the increase in TN could be attributed to N2-fixing behavior of Dolichospermum crassum (which reached high biomass and high heterocyte presence). In all other situations with high TN concentration, the cyanobacteria biomass was behaving as non-fixers (lack of presence of heterocytes), and hence increases in TN because of fixation would be unlikely.
In a temporal scale, the cyanobacteria blooms during the first summer period were three orders of magnitude higher than during the second summer. Also, cyanobacteria blooms during the first summer were found in the open water system of Laguna del Sauce (free of vegetation, field stations: SN, SS, and BM), whereas in the second summer the highest biomass occurred in Laguna del Potrero (dominated by submerged macrophytes) and Cisne.
Though the dominant heterocystous N2-fixing species showed similar biomass distributions along the TN and TP gradient, Dolichospermum never dominated at the same time than Aphanizomenon, Cuspidothrix, and Raphidiopsis. The latter three species did coexist during blooms. Indeed, the multivariate analysis indicates that these species displayed different responses to Secchi disc depth. This may relate to the fact that certain cyanobacteria bloom forming surface scums (e.g., Dolichospermum and Microcystis) while others bloom throughout the water column (e.g., Planktothrix and Limnothrix) (Chorus & Bartram, 1999; Paerl et al., 2016). Aphanizomenon and Cuspidothrix share similar morphology of their filaments; they are straight and have a very thin diameter (Komárek, 2013). Elongated forms tend to have high S:V ratio; high S:V ratio improves the access to resources (Lewis, 1976). Dolichospermum crassum, conversely, has coiled filament morphology, with higher cell size (Komárek, 2013). Coiling decreases the S:V of the organism (Kruk et al., 2010); low S:V decreases the access to resources (Lewis, 1976). Dolichospermum bloomed together with non-N-fixers of the Microcystis complex, which are also organisms with low S:V and which form cumulative blooms in the surface. Morphological differences may explain why Aphanizomenon, Cuspidothrix, and Raphidiopsis bloom together while Dolichospermum crassum do not. A recollection of the light traits (light efficiency, optimum irradiance, half saturation constant for light, growth rate) of cyanobacteria species will increase our ability to forecast which species will prevail under particular light conditions. Indeed, Schwaderer et al. (2011) showed that phytoplankton distributions along light gradients were linked to its light traits.
Final considerations
The different summer phytoplankton assemblages in Laguna del Sauce responded mostly to allogenic factors. The cyanobacteria component establishes whenever transparency was high (low turbidity and high Secchi disc depth), and temperature was high. The recurrence of heterocystous N2-fixing cyanobacteria blooms reflected the low average TN concentration and low TN:TP ratio in the system. The N2-fixing behavior is an adaptive response to low TN, though other morpho-functional traits may aid in predicting the species composition during blooms at high TN (e.g., light traits). These patterns can be used as an early warning tool for cyanobacterial bloom events, strengthening management measures to minimize interferences with the provision of drinking water.
References
Agawin, N. S. R., S. Rabouille, M. J. W. Veldhuis, S. Lidewij, S. Hol, H. M. J. van Overzee & J. Huisman, 2007. Competition and facilitation between unicellular nitrogen-fixing cyanobacteria and non-nitrogen-fixing phytoplankton species. Limnology and Oceanography 52: 2233–2248.
Azevedo, M. T. & C. L. Sant’Anna, 2003. Sphaerocavum, a new genus of planktic Cyanobacteria from continental water bodies in Brazil. Algological Studies 109: 79–92.
Carmichael, W. W. & G. J. Boyer, 2016. Health impacts from cyanobacteria algae blooms: implications for the North American Great Lakes. Harmful Algae 54: 194–2012.
Chorus, I. & J. Bartram, 1999. Toxic Cyanobacteria in water: a guide to their public health consequences, monitoring, and management, World Health Organization. E&FN Spon, London.
Crisci, C., R. Terra, J. P. Pacheco, B. Ghattas, M. Bidegain, G. Goyenola, J. J. Lagomarsino, G. Méndez & N. Mazzeo, 2017a. Multi-model approach to predict phytoplankton biomass and composition dynamics in a eutrophic shallow lake governed by extreme meteorological events. Ecological Modelling 360: 80–93.
Crisci, C., G. Goyenola, R. Terra, J. J. Lagomarsino, J. P. Pacheco, I. Díaz, L. González-Madina, P. Levrini, G. Méndez, M. Bidegain, B. Ghattas & N. Mazzeo, 2017b. Dinámica ecosistémica y calidad de agua: estrategias de monitoreo para la gestión de servicios asociados a Laguna del Sauce (Maldonado, Uruguay). INNOTEC 13: 46–57.
De Tezanos Pinto, P. & E. Litchman, 2010. Interactive effects of N:p ratios and light on nitrogen-fixer abundance. Oikos 119: 567–575.
Dolman, A. M., J. Rücker, F. R. Pick, J. Fastner, T. Rohrlack, U. Mischke & C. Widner, 2012. Cyanobacteria and Cyanotoxins: the Influence of Nitrogen versus Phosphorus. PLoS ONE 7: e38757.
Ferber, L. R., S. N. Levine, A. Lini & G. P. Livingston, 2004. Do cyanobacteria dominate in eutrophic lakes because they fix atmospheric nitrogen? Freshwater Biology 49: 690–708.
González-Madina, L., J. P. Pacheco, N. Mazzeo, P. Levrini, J. M. Clemente, J. J. Lagomarsino & C. Fosalba, 2017. Factores ambientales controladores del fitoplancton con énfasis en las cianobacterias potencialmente tóxicas en un lago somero utilizado como fuente de agua para potabilización: laguna del Sauce, Maldonado, Uruguay. INNOTEC 13: 26–35.
Guillard, R. L., 1978. Counting slides. In Sournia, A. (ed), Phytoplankton manual. UNESCO, Paris: 182–189.
Hillebrand, H., C. D. Durselen, D. Kirschtel, U. Pollingher & T. Zohary, 1999. Biovolume calculation for pelagic and benthic microalgae. Journal of Phycology 35: 403–424.
Inda, H. & M. Steffen, 2010. Bases técnicas para el manejo integrado de Laguna del Sauce y cuenca asociada. Universidad de la República y South American Institute for resilience and Sustainability Studies, Uruguay. ISBN 978-9974-0-0694-2.
INE 2011. Censo poblacional del Instituto Nacional de Estadística. http://www.ine.gub.uy/web/guest/censos-2011
Jacobsen, B. A. & P. Simonsen, 1993. Disturbance events affecting phytoplankton biomass, composition and species diversity in a shallow, eutrophic, temperate lake. Hydrobiologia 249: 9–14.
Komárek, J., 2013. Cyanoprokaryota 3. Teil/3rd Part: Heterocytous genera. In Büdel, B., G. Gärtner, L. Krienitz & M. Schagerl (eds), Süwasserflora von Mitteleuropa/Freshwater flora of Central Europe 19/3. Springer Spektrum, Berlin: 1130.
Komárek, J., J. Komárková-Legnerová & C. L. Sant’Anna, 2002. Two common Microcystis species (Chroococcales, Cyanobacteria) from tropical America, including M panniformis sp nov. Cryptogamie Algologie 23(2): 159–177.
Kruk, C., V. L. M. Huszar, E. Peeters, S. Bonilla, L. Costa, M. Lurling, C. S. Reynolds & M. Scheffer, 2010. A morphological classification capturing functional variation in phytoplankton. Freshwater Biology 55: 614–627.
Kumar, K., R. A. Mella-Herrera & J. W. Golden, 2010. Cyanobacterial Heterocysts. Cold Spring Harbor Perspectivein Biology 2: a00315.
Kützing, F.T., 1846. Tabulae phycologicae; oder, Abbildungen der Tange. Vol. I, fasc. 1 pp. 1-8, pls 1-10. Nordhausen: Gedruckt auf kosten des Verfassers In: commission bei W. Köhne.
Lemmermann, E., 1907. Algen I (Schizophyceen, Flagellaten, Peridineen). In Band, D. (ed), Kryptogamenflora der Mark Brandenburg und angrenzender Gebiete herausgegeben von dem Botanishcen Verein der Provinz Brandenburg. Verlag von Grebrüder Borntraeger, Leipzig: 1–304.
Lewis Jr., W. M., 1976. Surface/Volume ratio: implications for phytoplankton morphology. Science 28: 885–887.
Lund, J., C. Kipling & E. Le Cren, 1958. The inverted microscope method of estimating algal numbers and statistical basis of estimating by counting. Hydrobiologia 11: 143–170.
Margalef, R., 1978. Life forms of phytoplankton as survival alternatives in an unstable environment. Oceanologica Acta 1: 493–509.
Mazzeo, N., F. García-Rodríguez, A. Rodríguez, G. Méndez, C. Iglesias, H. Inda, G. Goyenola, S. García, C. Fosalba, S. Marroni, C. Crisci, L. Del Puerto, J. M. Clemente, J. P. Pacheco, C. Carballo, A. Kroeger, M. Vianna, M. Meerhoff, M. Steffen, J. J. Lagomarsino, M. Masdeu, N. Vidal, F. Texeira de Mello, I. González & D. Larrea, 2010. Estado trófico de Laguna del Sauce y respuestas asociadas. In Inda, H. & M. Steffen (eds), Bases técnicas para el manejo integrado de Laguna del Sauce y cuenca asociada. Montevideo, Uruguay: 31–49. ISBN 978-9974-0-0694-2.
Moisander, P. H., L. A. Cheshire, J. Braddy, E. S. Calandrino, M. Hoffman, M. F. Piehler & H. W. Paerl, 2012. Facultative diazotrophy increases Cylindrospermopsis raciborskii competitiveness under fluctuating nitrogen availability. FEMS Microbial Ecology 79(3): 800–811.
Mugidde, R., R. E. Hecky, L. L. Hendzel & W. D. Taylor, 2003. Pelagic nitrogen fixation in lake Victoria (East Africa). Journal of Great Lakes Research 29: 76–88.
Muller, R. & O. Wiedemann, 1955. Die Bestimmung des Nitrations in Wasser. Von Wasser 22: 247–271.
Murphy, J. & J. P. Rilley, 1962. A modified single-solution method for the determination of phosphate in natural waters. Analytica Chimica Acta 27: 31–36.
Nusch, E. A., 1980. Comparison of different methods for chlorophyll and pheopigment determination. Archives of Hydrobiology 14: 14–36.
OECD, 1982. Eutrophication of Waters, Monitoring, Assessment and Control. Organisation for Economic Cooperation and Development in Paris, Paris.
Pacheco, J. P., C. Iglesias, M. Meerhoff, C. Fosalba, G. Goyenola, F. Teixeira-de Mello, S. García, M. Gelós & F. García-Rodríguez, 2010. Phytoplankton community structure in five subtropical shallow lakes with different trophic status (Uruguay): a morphology-based approach. Hydrobiologia 646(1): 187–197.
Padisák, J., 1992. Seasonal succession of phytoplankton in a large shallow lake (Balaton, Hungary)—a dynamic approach to ecological memory, its possible rol and mechanisms. Journal of ecology 80: 217–230.
Padisák, J. & V. Istvánovics, 1997. Differential response of blue-green algal groups to phosphorus load reduction in a large shallow lake: Balaton, Hungary. Verhandlungen des Internationalen Verein Limnologie 26: 574–580.
Paerl, H. W., W. S. Gardner, K. E. Havens, A. R. Joyner, M. J. McCarthy, S. E. Newell & J. T. Scott, 2016. Mitigating cyanobacterial harmful algal blooms in aquatic ecosystems impacted by climate change and anthropogenic nutrients. Harmful Algae 54: 213–222.
R Core Team (2016). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/.
Rajaniemi, P., J. Komárek, L. Hoffmann, P. Horuzek, K. Kastocská & K. Sivonen, 2005. Taxonomic consequences from the combined molecular phenotype evaluation of selected Anabaena and Aphanizomenon strains. Algological Studies 111: 371–391.
Reynolds, C. S., 2006. Ecology of phytoplankton. Cambridge University Press, Cambridge.
Reynolds, C. S., V. Huszar, C. Kruk, F. L. Naselli & S. Melo, 2002. Towards a functional classification of the freshwater phytoplankton. Journal of Plankton Research 24: 417–428.
Rodríguez, A., G. Méndez, S. Kausas, J. M. Clemente, A. Kroger & N. Mazzeo, 2010. Importancia de la carga externa e interna de nutrientes en el estado trófico de la Laguna del Sauce. In Inda, H. & M. Steffen (eds), Bases técnicas para el manejo integrado de Laguna del Sauce y cuenca asociada. Montevideo, Uruguay: 53–61.
Schwaderer, A., K. Yoshiyama, P. de Tezanos Pinto, N. G. Swenson, C. A. Klausmeier & E. Litchman, 2011. Eco-evolutionary differences in light utilization traits and distributions of freshwater phytoplankton. Limnology and Oceanography 56: 589–598.
Seenayya, G., Subba Raja, N. (1972). On the ecology and systematic of the alga known as Anabaenopsis raciborskii (Wolosz.) Elenk. And a critical evaluation of the forms described under the genus Anabaenopsis. In: Desikachary, T.V. (ed) Papers submitted to the First International Symposium on Taxonomy and Biology of blue-green algae. pp. 52–57.
Uhelinger, V., 1964. Étude statistique des méthodes de dénobrement planctonique. Archive de Science 17: 121–223.
Utermöhl, H., 1985. Zur Vervolkommung der quantitativen Phytoplankton-Methodik. Mitteilungen der Internationale Vereiningung fuer Theoretische and Angewandte Limnologie 9: 1–38.
Valderrama, J. C., 1981. The simultaneous analysis of total N and total P in natural waters. Marine Chemistry 10: 1009–1022.
Vitousek, P. M., et al., 2002. Towards an ecological understanding of biological nitrogen fixation. Biogeochemistry 57: 1–45.
Weithoff, G., 2003. The concepts of plant functional types and functional diversity in lake phytoplankton: a new understanding of phytoplankton ecology? Freshwater Biology 48: 1669–1675.
Wolk, P. C., A. Ernst & J. Elahi, 1994. Heterocyst metabolism and development. In Bryant, D. A. (ed), The Molecular Biology of Cyanobacteria. Kluwer Academic Publishers, Dordrecht: 769–823.
Yema, L., E. Litchman & P. de Tezanos Pinto, 2016. The role of heterocytes in the physiology and ecology of bloom forming harmful cyanobacteria. Harmful Algae 60: 131–138.
Zhang, C. C., S. Laurent, S. Sakr, L. Peng & S. Bédu, 2006. Heterocyst differentiation and pattern formation in cyanobacteria: A chorus of signals. Molecular Microbiology 59: 367–375.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: S. Nandini, S.S.S. Sarma, Erik Jeppesen & Linda May / Shallow Lakes Research: Advances and Perspectives
Rights and permissions
About this article

Cite this article
González-Madina, L., Pacheco, J.P., Yema, L. et al. Drivers of cyanobacteria dominance, composition and nitrogen fixing behavior in a shallow lake with alternative regimes in time and space, Laguna del Sauce (Maldonado, Uruguay). Hydrobiologia 829, 61–76 (2019). https://doi.org/10.1007/s10750-018-3628-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-018-3628-6