
Abstract
Malaria infection is a multifactorial disease partly modulated by host immuno-genetic factors. Recent evidence has demonstrated the importance of Interleukin-17 family proinflammatory cytokines and their genetic variants in host immunity. However, limited knowledge exists about their role in parasitic infections such as malaria. We aimed to investigate IL-17A serum levels in patients with severe and uncomplicated malaria and gene polymorphism’s influence on the IL-17A serum levels. In this research, 125 severe (SM) and uncomplicated (UM) malaria patients and 48 free malaria controls were enrolled. IL-17A serum levels were measured with ELISA. PCR and DNA sequencing were used to assess host genetic polymorphisms in IL-17A. We performed a multivariate regression to estimate the impact of human IL-17A variants on IL-17A serum levels and malaria outcomes. Elevated serum IL-17A levels accompanied by increased parasitemia were found in SM patients compared to UM and controls (P < 0.0001). Also, the IL-17A levels were lower in SM patients who were deceased than in those who survived. In addition, the minor allele frequencies (MAF) of two IL-17A polymorphisms (rs3819024 and rs3748067) were more prevalent in SM patients than UM patients, indicating an essential role in SM. Interestingly, the heterozygous rs8193038 AG genotype was significantly associated with higher levels of IL-17A than the homozygous wild type (AA). According to our results, it can be concluded that the IL-17A gene rs8193038 polymorphism significantly affects IL-17A gene expression. Our results fill a gap in the implication of IL-17A gene polymorphisms on the cytokine level in a malaria cohort. IL-17A gene polymorphisms also may influence cytokine production in response to Plasmodium infections and may contribute to the hyperinflammatory responses during severe malaria outcomes.
Similar content being viewed by others

Background
Malaria is an infectious disease, potentially fatal, caused by Plasmodium protozoan parasites and transmitted by female Anopheles mosquitoes. According to the WHO, malaria is one of the leading causes of death worldwide. Globally, there were an estimated 247 million malaria cases in 2021 in 84 malaria-endemic countries, increasing from 245 million in 2020, with most of this increase coming from countries in the African Region. Furthermore, in 2020, malaria deaths increased by 10% compared with 2019, and between 2019 and 2021, there were 63,000 deaths due to disruptions to essential malaria services during the COVID-19 pandemic. Thus, malaria remains a major public health problem, especially in Africa [1]. In Senegal, Malaria is endemic throughout, and the entire population is exposed to the disease. In 2020, 0.7% of malaria deaths worldwide occurred in Senegal. The number of malaria cases fell by 4.4% between 2017 and 2020, from 52 to 50 per 1,000 inhabitants at risk, while the number of malaria deaths rose slightly by 1.8% over the same period, from 0.24 to 0.245 per 1,000 inhabitants at risk.
The malaria pathogenesis is complex and needs to be elucidated. During blood-stage infection, the host’s immune system produces proinflammatory cytokines to eliminate the parasite, including IL-6, IFN-γ, and TNF, which are pivotal in controlling the parasite’s growth and elimination. In many studies, the high levels of some pro-inflammatory cytokines have been protective in malaria [2,3,4]. Pro-inflammatory biomarkers were more elevated in cerebral malaria than in non-cerebral malaria patients [5]. Regulatory cytokines such as transforming growth factor-β (TGF-β) and IL-10 balance the pro-inflammatory and anti-inflammatory responses. However, in many cases, cytokines have a double role. On the one hand, they contribute to parasitic clearance; on the other, they are responsible for pathological changes encountered in malaria. Cytokine-modulating strategies may represent a promising modern approach to disease management [3], and the Circulating levels of cytokines have the potential to be biomarkers for severity or protection against malaria.
Recently, the IL-17 proinflammatory cytokine has gained attention among malaria researchers because of its protective role in immunity against extracellular pathogens [6,7,8] and for the clearance of intracellular pathogens [9,10,11]. In addition to its essential role in protective immunity, IL-17 is critical in the pathogenesis of various autoimmune inflammatory diseases. IL-17 is a cytokine family that plays a vital role in innate and adaptive immune systems [12,13,14,15]. The IL-17 gene is located on chromosome 6p12, comprises three exons and two introns and is coded with six protein members (IL-17A-F). IL-17A is the most essential member of the IL-17 family. The IL-17 receptor family now comprises 5 members (IL-17RA, RB, RC, RD and RE) [16,17,18].
In mice, it has been demonstrated that elevated IL-17 levels and high IL-4, IL-12α and IFN-γ levels may be a marker of protection against Plasmodium berghei [19]. However, the role of IL-17 in human malarial infection outcomes is poorly described, even if increased IL-17 levels in vivax and falciparum malaria and disease severity have been reported [5, 20]. Further studies are needed to evaluate the implication of IL-17 cytokines' circulation levels and association in malaria protection and/or pathogenesis.
Single Nucleotide Polymorphisms (SNPs) in encoding regions of the IL-17A gene may influence changes in its expression and, potentially, malaria pathogenesis and risk. IL-17A SNPs have been linked to several malignancies, including gastric and breast cancer [21, 22]. However little is known about the association between IL-17 gene variation and malaria.
To understand the specific IL-17A roles in malaria infection, we conducted this work to analyse IL-17A levels in a retrospective Senegalese cohort, including healthy controls and severe and uncomplicated malaria subjects. Then, the IL-17A gene and its flanking regions were sequenced in individual samples. The SNPs were analysed among individuals concerning malaria disease status to detect their influence on the IL-17A serum levels and its potential associations with malaria severity.
Methods
Study participants
Our study was conducted in Senegal, a malaria-endemic country in the Sahelian zone of West Africa. Malaria patients were enrolled using a retrospective cohort from the Hospital Principal of Dakar between the 2012–2015 period and corresponded to subjects with Plasmodium-positive Quantitative Buffy Coat (QBC) [23, 24]. These patients were classified into Uncomplicated Malaria (UM) and Severe Malaria (SM) groups, according to the criteria defined by Saissy et al. 2003 and previously described in [25]. The inclusion criteria for these patients were: firstly, black Senegalese individuals with P. falciparum infection confirmed in diagnosis; secondly, persons born in Senegal whose parents and grandparents were born in Senegal; and finally, individuals who have not travelled in the last three months of their hospital admission. The healthy control subjects corresponded to the exposed and uninfected subjects group in the same areas and inclusion criteria. They correspond to patients visiting the hospital for annual health check-up with a Plasmodium-negative QBC test.
The study protocol was approved by UCAD’s Committee on Research and Ethics (CER), which considers that the research proposed respects the appropriate ethical standard and, as a result, approves its execution under “Protocole 0344/2018/CER-UCAD”. Written informed consent was obtained from adult participants and parents or legal representatives of children.
The study included 48 CTR, 54 UM and 71 SM subjects. The study’s objectives have been explained clearly using the local dialect before including patients in hospital centres.
Serum collection and IL-17A quantification
Blood samples were collected from malaria patients and controls and drawn into EDTA vacutainer tubes. Samples were centrifuged, plasma aliquoted, and stored at − 20 °C until testing. Sandwich quantified serum levels of IL-17A enzyme-linked immunosorbent assays (ELISA) using the pre-designed kit (Elabscience Biotechnology ®, USA) as per the manufacturer. The sensitivity of the assay protocol was 2.38 pg/mL, and the coefficient of variation (CV) for intra-assays and inter-assays was < 8% and < 10%, respectively. The micro ELISA plates provided in the kit have been pre-coated with an antibody specific to Human IL-17A. Standards and samples were added to the micro ELISA plate wells and combined with the specific antibody. Then a biotinylated detection antibody specific for Human IL-17A and Avidin-Horseradish Peroxidase (HRP) conjugate were added successively to each microplate well and incubated. The wells were washed 3 times with 350 uL of wash buffer, and the substrate solution was added to each well. Only those wells that contain Human IL-17A, biotinylated detection antibody and Avidin-HRP conjugate will have appeared blue in colour. The enzyme–substrate reaction was terminated by adding a stop solution, and the colour turned yellow. The optical density (OD) was measured spectrophotometrically at a wavelength of 450 nm ± 2 nm. The OD value was proportional to the concentration of Human IL-17A. We have calculated the concentration of Human IL-17A in the samples by comparing the OD of the samples to the standard curve. The sensitivity of the assay protocol was 18.75 pg/mL, and the coefficient of variation (CV) for intra-assays and inter-assays was < 10%.
Genotyping
DNA was extracted from the peripheral blood of each subject by using standard Qiagen Kits according to the manufacturer’s recommendations. The concentration and quality of the extracted DNA were measured using a NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). The IL-17A polymorphisms were genotyped using the polymerase chain reaction method. The Oligonucleotide primers used to amplify promoters and exon regions are listed in Table 1. The PCR reactions were performed using a Gotaq®Green Master Mix (Promega, Germany) in a total volume of 25 µl containing 25 ng of genomic DNA (5 ng/µl) and 2.5 µL of each primer (10 μM). The PCR conditions were initial denaturation at 95 °C for 5 min, 35 cycles at 95 °C for 30 s, 60 °C for 30 s, and 72 °C for 5 min, with a final extension at 72 °C for 10 min. The amplicons were purified using BioGel P100 gels (Bio-Rad). Sequencing reactions (2 µL of PCR product) were performed using the dye terminator v3.1 method in an ABI PRISMs 3730 DNA Analyzer (Applied Biosystems, Foster City, CA, USA). Sequencing conditions were: 96 °C for 5 min, 25 cycles of 96 °C for 10 s, 60 °C for 4 min and 15 °C forever, and PCR products were purified with Sephadex G50 superfine columns (GE Healthcare). Alignment of acquired sequences and SNP discovery were performed using NC_000006.12 as a reference. Analysis was performed with Genalys version 2.0b software [26].
Statistical analysis
GraphPad Prism v9.5.1 was employed for all statistical analyses. One-way variance analysis ANOVA or Student statistical tests compared serum levels of IL-17A in malaria groups and controls. We calculated Allelic frequencies and Hardy–Weinberg equilibrium, as described previously (Rodriguez, Gaunt & Day, 2009) [27]. Then, as reported previously, the differences in allelic frequencies between the three groups (SM, UM, CTR) were determined using the logistic regression analysis method [28, 29]. Next, associations between IL-17A genotypes and serum levels and malaria outcomes were performed using the Mann–Whitney test, and then associations with P values < 0.05 were considered statistically significant.
Results
Characteristics of malaria patients and healthy controls
A total of 125 malaria patients and 48 controls were included in this retrospective study. The clinical characteristics of the malaria and control groups are summarized in Table 2. The mean values of all haematological parameters included in our study were determined in three categories of enrolled subjects, and the statistical analysis was done using an ANOVA test. A significant difference was observed in parameters while comparing malaria cases and controls, such as age, haemoglobin level, blood cell parameters and leukocyte cells. However, there were no statistically significant differences in monocytes, Basophils. We then compared the same parameters between severe and uncomplicated malaria patients using the Student's t-test. We found the same results as above except for lymphocytes and neutrophils, for which the differences in means between SM and UM were insignificant.
Severe malaria patients displayed higher serum IL‑17A compared to uncomplicated malaria and controls
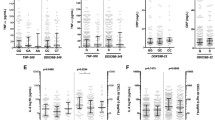
ELISA test quantified the level of IL-17A. As shown in Fig. 1, the mean level of IL-17A was significantly higher in the SM group (mean ± SE: 37.74 ± 140 pg/mL) compared to that of the UM and control groups (mean ± SD:1.69 ± 2.4 pg/ml and 0.67 ± 1.07 pg/mL, respectively; P < 0.001) (Fig. 1a). In the SM group, we observed a lower level of IL-17A in patients who were deceased compared to those who survived (mean ± SD: 25.92 ± 46.48 and 40.78 ± 150.67 pg/mL, respectively). Still, the difference was not significant (P = 0.56); the lack of significance may be partly due to the small sample size (Fig. 1b). The parasitemia at the time of diagnosis in SM patients was higher than that of the UM patients (mean ± SD: 9039.54 ± 17891 and 3084 ± 14092, respectively; P = 0.0022). In addition, there was a decrease in parasitemia in the deceased compared to the survivors, but the difference was not significant (mean ± SD: 9915.54 ± 19185 and 6496 ± 7161, respectively; P = 0.8) (Fig. 1c, d).

Parasite density and IL17A levels in different categories of enrolled subjects. a, b Serum level IL-17 between SM, UM and Control group and survivors and deceased SM patients. c, d Parasitemia between SM, UM and Control group, survivors, and deceased SM patients. Data representing mean pg/ml ± SE for IL17A and mean P/uL for parasitemia were analysed with one way ANOVA or Student Test for comparison. P < 0.05 was considered statistically significant
Distribution of IL-17A polymorphisms and association with the risk of severe malaria outcome
We analyzed genetic variations on the IL-17A gene in the promoter region and all the coding regions by sequencing and identified eight SNPs, including 4 SNPs located in the 5’UTR: IL-17A + 521A/C (rs9791323), IL-17A + 606A/G (rs3819024), IL-17A + 849G/A (rs2275913), IL-17A + 973G/A (rs8193037); two SNP located within intron 1: IL-17A + 1090G/A (rs3819025), IL-17A + 1198A/G (rs8193038); one located within exon 3: IL-17A + 3840G/A (rs17880588) and one located in the 3′UTR: IL-17A + 5151C/T (rs3748067) (Fig. 2).

Mapping of Interleukin -17A (IL-17A) gene and the polymorphisms genotyped. Schema of the gene encoding Interleukin 17 A (IL17A) (a) The human IL17A gene (located at 6p12-2) contains 3 exons that span 4 kb. Exons are described by number. AUG: the transcription start site. Black arrows mark eight specific polymorphisms identified in the IL-17A gene
The Minor Allele frequencies (MAF) of the eight SNPs loci of the IL-17A gene were identified using the Hardy–Weinberg equilibrium test (Table 3). Among them, 6 SNP (rs9791323, rs3819024, rs2275913, rs3819025, rs17880588 and rs3748067) were detected with high frequencies (with MAF > 3%), unlike 2 other SNPs (rs8193037 and rs8193038) with MAF < 3% were observed (Table 3). First, comparisons were performed among the three groups to test whether polymorphisms were associated with malaria severity. Then, statistical IL-17A polymorphisms analysis was performed using logistic regression tests with an adjustment for potential confounders such as Hb polymorphisms. The SNPS rs3819024 and rs3748067 yielded a significant association with Severe Malaria. For SM vs UM, the P = 0.007 (OR 2.61, 95% CI 0.35– 0.91) and P = 0.04 (OR 0.32, 95% CI 1–2.1), respectively. In addition, the SNP rs9791323 was associated with uncomplicated malaria P = 0.045 (OR 3.78, 95% CI 2.13 – 7.8) with higher MAF in UM (11.3%) patients compared to controls (3.3%).
Association of SNP IL-17A + 1198 A/G (rs8193038)) and serum IL-17A concentration
The relationship between the IL-17A polymorphisms and serum IL-17A concentration was analysed regardless of the differences in the study groups. We compared IL17A levels between the different genotypes of the polymorphisms analysed in the study and found differences in levels depending on genotype. We found that some genotypes were associated with increased IL17A levels (Fig. 3c, f, h), while the opposite was true for others (Fig. 3a, b, d, e, g).

Concentration of IL-17A according to its genotypes. ELISA quantified serum levels of IL-17A in a total of 173 subjects comprising Severe and Uncomplicated malaria subjects and Controls. Mean IL-17A levels in the different genotypes of rs2275913 (a) rs3819024 (b) rs2275913 (c) rs81933037 (d) rs3819025 (e) rs8193038 (f) rs17880588 (g) rs3748067 (h) Polymorphisms were compared by ANOVA followed by Tukey’s post-test. No significant association between IL-23A (rs11171806) and levels of IL-23 was observed (b). A p-value of less than 0.05 was taken as significant
However, only the heterozygous rs8193038 AG genotype is significantly associated with higher levels of IL-17A amongst the whole study groups compared to the homozygous rs8193038 AA genotype (OR = 4.9, 95% CI = (2.01- 8.13), P< 0.001).
Discussion
Analysis of the genetic effects of inflammatory response gene variants such as inflammatory cytokines is a key step in malaria research to understand the underlying mechanisms of pathogenesis. This knowledge will be essential for identifying specific therapies to prevent mortality or adverse complications associated with severe malaria and the long-term consequences that represent a heavy burden in endemic regions [30]. It is understood that cytokine gene polymorphisms could affect the serum levels of cytokines by influencing transcriptional regulation. The IL-17 cytokine family is a relatively new family linked to adaptive and innate immune systems. IL-17A are members of the IL-17 cytokine family, essential for the pathogenic activity of IL-17 cells and the production of various proinflammatory mediators in the body [9, 31]. IL-17A is a multifunctional cytokine which has a protective role in immunity for the clearance of intracellular pathogens such as Plasmodium spp. [9,10,11, 19, 32, 33], making it a good therapeutic biomarker in malaria diseases. However, the role of IL-17A in malaria has not yet been extensively investigated. In addition, the association of common polymorphisms with malaria predisposition and cytokine levels was never analysed in Senegal.
Thus, the present study explored IL-17A cytokine levels and gene polymorphism’s influence on IL-17A serum levels. In recent years, evaluating SNPs has been considered a common approach for testing the impact of human genetic variation on diseases [34].
We have determined the serum IL-17A levels and genotyped IL-17A variants in Senegalese severe and uncomplicated malaria patients and controls. We observed elevated IL-17A levels in SM patients compared to the UM and healthy cases. Moreover, the high parasitemia in the SM group accompanied the IL-17A increase. This indicates that IL-17A has an essential regulatory role in malaria infection, controlling the intensity of the immune response, as described in the experimental model, as well as human malaria and several other infectious diseases [5, 19, 35, 36]. IL-17 production is associated with a very high occurrence of chronic inflammation and immunopathological conditions [31]. Recent data suggest that IL-17 contributes to host protection against diverse infectious organisms during sepsis while inducing hyperinflammation with detrimental outcomes for the host under certain conditions [37]. Earlier investigations in the experimental model have deciphered the essential role of IL-17. In P. vivax infection, authors suggest that increasing serum IL-17 levels in malaria patients could be considered a host adaptation mechanism to control changes in blood viscosity, and IL-17 could thus be used as an immunomodulatory agent [38]. IL-17 appears to act on erythrocytes by remodelling their cell membrane; it is well-known that erythrocytes in malaria are very sensitive to osmotic shock [38].
We found an elevated level of IL-17A in severe malaria patients who were survivors compared to those who were deceased. Our results seem to confirm the results of Helegbe et al., which showed elevated IL-17 levels together with high IL-4, IL-12α, and IFN-γ levels may be a marker of protection, and the mechanism may be controlled by host factors [19]. Thus, pro-inflammatory IL-17A cytokine seems to have been protective against fatal malaria. Furthermore, the data agree with the observations of Oyegue-Liabagui et al. [20], who noted a correlation between Th17 cell count and overall survival in patients with malaria in children.
Immuno-genetic variants are associated with diverse degrees of malaria susceptibility, including cytokine gene polymorphisms that modify their expression and circulating protein levels to reflect inflammatory or anti-inflammatory responses [39,40,41]. Polymorphisms in the IL-17A cytokine can impact the activity and expression of inflammatory mediators, which can affect interleukin-17 activity [42, 43]. IL-17A gene polymorphisms have been linked to several malignancies, including gastric and breast cancer [21, 22]. However little is known about the association between IL-17 gene variation and malaria. In this study, we performed a genetic analysis of the variations of the IL-17A gene. We identified 8 SNPs in the IL-17A gene; among them, 6 SNPs (rs9791323, rs3819024, rs2275913, rs3819025, rs17880588 and rs3748067) were detected with high frequencies (with MAF > 3%) at opposite to 2 other SNPs (rs8193037 and rs8193038) with MAF < 3% were observed. Then, statistical IL-17A polymorphism analysis was performed using logistic regression to test whether polymorphisms were associated with malaria severity. For the first time, we identified 2 SNPs associated with severe malaria and one associated with uncomplicated malaria. We found that the SNP rs3748067 reduced the risk of severe malaria (odds ratio (OR) = 0.32; P = 0.04). Instead, the SNP rs3819024 was associated with an increased risk of severe malaria (odds ratio (OR) = 2.61; P = 0.007). Then, the SNP rs9791323 was associated with a high risk of uncomplicated malaria (odds ratio (OR) = 3.78; P = 0.045). We IL-17A rs3819024 G could also be considered a biomarker of malaria severity.
Our data reinforce our knowledge of the genetic variants of the IL-17 cytokine family and their potential roles in malaria. Even though the involvement of IL-17A variants has yet to be fully elucidated, a previous study had shown that IL-17F (rs6913472 and rs4715291) and IL-17RA (rs12159217 and rs41396547) polymorphisms independently modulate susceptibility to Cerebral Malaria and provide evidence that IL-17F protects against it [44]. The role of SNPs in malaria disease and immunological disorders has been previously reported [45,46,47]. Associations between cytokine polymorphisms and malaria support that cytokine gene polymorphisms have an unquestionable role in the orchestration of the immune response, leading to different functional scenarios, which in turn influence the outcome of malaria disease establishment and evolution [20, 48, 49].
The relationship between the IL-17A polymorphisms and serum IL-17A concentration was analysed. The SNPs rs3748067, rs3819024 and rs9791323 did not show an association with malaria outcome and IL-17A level. Interestingly, we found that the heterozygous rs8193038 AG genotype is significantly associated with higher levels of IL-17A amongst the whole study groups compared to the homozygous rs8193038 AA genotype (OR = 4.9, 95% CI = (2.01- 8.13), P < 0.001). This data suggests that rs8193038 polymorphism significantly affects IL-17A gene expression. However, this SNP is located at the intron 1 region, which could correspond to a splicing site and explain our result [50, 51]. It is well known that the introns regulate gene expression; they contain enhancers or other cis-acting elements that promote the initiation or elongation of transcription. Introns are also involved in alternative splicing and genome imprinting. Intron splicing increases mRNA stability in the nucleus [52]. It has been shown that intronal SNPs regulate protein synthesis by mRNA splicing [53]. In addition, functional SNPs in introns are sometimes linked to SNPs in neighbouring genes, influencing mRNA splicing, among other things [54]. The genome-wide analysis of human SNPs near splice sites revealed 1300 SNPs, which are probably capable of modifying the protein by changing splicing [55]. Further studies are needed to elucidate how this intronic rs8193038 SNP influences mRNA splicing and IL-17A expression.
Our results fill a gap in the implication of IL-17A gene polymorphisms on the cytokine level in a Senegalese malaria cohort. IL-17A gene polymorphisms also may influence cytokine production in response to Plasmodium infections and may be contribute to the hyperinflammatory responses during severe malaria outcomes. A series of studies performed in the last decade emphasized the IL-17A SNPs, particularly the rs2275913 variant, and serum cytokines levels in numerous pathogenesis. A recent study by Lang et al. revealed the association between the IL-17A rs2275913 variant with higher cytokine serum levels and predisposed Preeclampsia development in Chinese patients [56]. In leprosy, Farag et al. have demonstrated that IL17A rs2275913 genotype GG was associated with significantly increased IL-17A levels in Egyptian patients [57]. Finally, another study by Li et al. revealed that IL-17A polymorphisms may influence hepatocellular carcinoma risk in chronic hepatitis B virus infection via regulating IL-17A production [58]. However, our study found no association between rs2275913 variant and serum IL-17A levels, even if a high serum level of IL-17A was associated with the heterozygous rs2275913 GA genotype. The lack of significance may be due to a limited number of subjects. These relationships could likely reach statistical significance in a larger cohort of patients. Our results suggest that the association between IL-17 gene polymorphisms and serum levels may depend on ethnic group populations and/or pathogenesis mechanisms. Then, we found that different IL-17A alleles play different roles in immunity, producing different cytokine levels and disease outcomes, as reported in other studies [59].
Conclusions
The current report revealed an elevated level of IL-17A in severe malaria patients compared to healthy controls in Senegalese cohort. Furthermore, heterozygous mutant and minor alleles of IL-17A rs3819024 and rs3748067 polymorphisms predisposed subjects for the development of SM. Interestingly, the current report further validated the functional relevance of IL-17A (rs8193038) variants and demonstrated the association of mutants with elevated IL-17A levels. However, further studies, including more significant sample sizes in the different populations, are required to validate the observations of the present study. In addition, further investigation on the role of IL-17 and its interplay with other immune factors needs to be conducted in clinical settings. In the future, we plan to carry out a mechanistic study to understand the role of these IL-17A SNPs in gene expression. We intend to measure the expression of IL-17A mRNA to confirm our results. We will use a genome-wide association study to screen the IL-17A gene better and characterize the position of the SNPs better. Finally, we will perform proteomic studies and use bioinformatics tools to understand the molecular function of SNPs.
Availability of data and materials
The data and materials supporting the conclusions of the study are available from the corresponding author on request.
Abbreviations
- COVID:
-
Coronavirus disease 2019
- CTR:
-
Control
- EDTA:
-
Ethylene Diamine Tetraacetic Acid
- ELISA:
-
Enzyme-Linked Immuno Assay
- IFN:
-
Interferon
- IL:
-
Interleukin
- MAF:
-
Minor Allele Frequency
- PCR:
-
Polymerase Chain Reaction
- SM:
-
Severe Malaria
- SNP:
-
Single Nucleotide Polymorphism
- TGF:
-
Tumor Growth Factor
- TNF:
-
Tumor Necrosis Factor
- UCAD:
-
Universite Cheikh Anta Diop
- UM:
-
Uncomplicated Malaria
- WHO:
-
World Health Organization
References
World Health Organization. World malaria report 2020. Geneva: WHO; 2021. p. 1–178.
Obeng-Aboagye E, Frimpong A, Amponsah JA, Danso SE, Owusu EDA, Ofori MF. Inflammatory cytokines as potential biomarkers for early diagnosis of severe malaria in children in Ghana. Malar J. 2023;22(1):220.
Popa GL, Popa MI. Recent advances in understanding the inflammatory response in malaria: a review of the dual role of cytokines. J Immunol Res. 2021;2021:7785180.
Angulo I, Fresno M. Cytokines in the pathogenesis of and protection against malaria. Clin Diagn Lab Immunol. 2002;9(6):1145–52.
Dieye Y, Mbengue B, Dagamajalu S, Fall MM, Loke MF, Nguer CM, Thiam A, Vadivelu J, Dieye A. Cytokine response during non-cerebral and cerebral malaria: evidence of a failure to control inflammation as a cause of death in African adults. PeerJ. 2016;4:e1965.
Agak GW, Mouton A, Teles RM, Weston T, Morselli M, Andrade PR, Pellegrini M, Modlin RL. Extracellular traps released by antimicrobial TH17 cells contribute to host defense. J Clin Invest. 2021;131(2):e141594.
Puerta-Arias JD, Mejia SP, Gonzalez A. The role of the interleukin-17 axis and neutrophils in the pathogenesis of endemic and systemic mycoses. Front Cell Infect Microbiol. 2020;10:595301.
Dixon B, Lee TJ, Contreras Healey DC, Li J, Goettel JA, Piazuelo MB, Algood HMS. IL-17 receptor signaling through IL-17A or IL-17F is sufficient to maintain innate response and control of helicobacter pylori immunopathogenesis. Immunohorizons. 2022;6(2):116–29.
Evans HG, Gullick NJ, Kelly S, Pitzalis C, Lord GM, Kirkham BW, Taams LS. In vivo activated monocytes from the site of inflammation in humans specifically promote Th17 responses. Proc Natl Acad Sci U S A. 2009;106(15):6232–7.
Banerjee A, Bhattacharya P, Joshi AB, Ismail N, Dey R, Nakhasi HL. Role of pro-inflammatory cytokine IL-17 in Leishmania pathogenesis and in protective immunity by Leishmania vaccines. Cell Immunol. 2016;309:37–41.
Hansakon A, Ngamskulrungroj P, Angkasekwinai P. Contribution of Laccase Expression to Immune Response against Cryptococcus gattii Infection. Infect Immun. 2020;88(3):e00712.
Bunte K, Beikler T. Th17 Cells and the IL-23/IL-17 axis in the pathogenesis of periodontitis and immune-mediated inflammatory diseases. Int J Mol Sci. 2019;20(14):3394.
Nadeem A, Al-Harbi NO, Alfardan AS, Ahmad SF, AlAsmari AF, Al-Harbi MM. IL-17A-induced neutrophilic airway inflammation is mediated by oxidant-antioxidant imbalance and inflammatory cytokines in mice. Biomed Pharmacother. 2018;107:1196–204.
Schon MP, Erpenbeck L. The interleukin-23/Interleukin-17 axis links adaptive and innate immunity in psoriasis. Front Immunol. 2018;9:1323.
Li G, Zhang Y, Qian Y, Zhang H, Guo S, Sunagawa M, Hisamitsu T, Liu Y. Interleukin-17A promotes rheumatoid arthritis synoviocytes migration and invasion under hypoxia by increasing MMP2 and MMP9 expression through NF-kappaB/HIF-1alpha pathway. Mol Immunol. 2013;53(3):227–36.
Kolls JK, Linden A. Interleukin-17 family members and inflammation. Immunity. 2004;21(4):467–76.
Kawaguchi M, Adachi M, Oda N, Kokubu F, Huang SK. IL-17 cytokine family. J Allergy Clin Immunol. 2004;114(6):1265–73. Quiz 1274.
McGeachy MJ, Cua DJ, Gaffen SL. The IL-17 family of cytokines in health and disease. Immunity. 2019;50(4):892–906.
Helegbe GK, Huy NT, Yanagi T, Shuaibu MN, Kikuchi M, Cherif MS, Hirayama K. Elevated IL-17 levels in semi-immune anaemic mice infected with Plasmodium berghei ANKA. Malar J. 2018;17(1):169.
Oyegue-Liabagui SL, Bouopda-Tuedom AG, Kouna LC, Maghendji-Nzondo S, Nzoughe H, Tchitoula-Makaya N, Pegha-Moukandja I, Lekana-Douki JB. Pro- and anti-inflammatory cytokines in children with malaria in Franceville. Gabon Am J Clin Exp Immunol. 2017;6(2):9–20.
Long ZW, Yu HM, Wang YN, Liu D, Chen YZ, Zhao YX, Bai L. Association of IL-17 polymorphisms with gastric cancer risk in Asian populations. World J Gastroenterol. 2015;21(18):5707–18.
Wang L, Jiang Y, Zhang Y, Wang Y, Huang S, Wang Z, Tian B, Yang Y, Jiang W, Pang D. Association analysis of IL-17A and IL-17F polymorphisms in Chinese Han women with breast cancer. PLoS One. 2012;7(3):e34400.
Thiam F, Diop G, Coulonges C, Derbois C, Mbengue B, Thiam A, Nguer CM, Zagury JF, Deleuze JF, Dieye A. G6PD and HBB polymorphisms in the Senegalese population: prevalence, correlation with clinical malaria. PeerJ. 2022;10:e13487.
Diop G, Derbois C, Loucoubar C, Mbengue B, Ndao BN, Thiam F, Thiam A, Ndiaye R, Dieye Y, Olaso R, et al. Genetic variants of RNASE3 (ECP) and susceptibility to severe malaria in Senegalese population. Malar J. 2018;17(1):61.
Saissy JM, Rouvin B, Koulmann P. Severe malaria in intensive care units in 2003. Med Trop (Mars). 2003;63(3):258–66.
Takahashi M, Matsuda F, Margetic N, Lathrop M. Automated identification of single nucleotide polymorphisms from sequencing data. J Bioinform Comput Biol. 2003;1(2):253–65.
Rodriguez S, Gaunt TR, Day IN. Hardy-Weinberg equilibrium testing of biological ascertainment for Mendelian randomization studies. Am J Epidemiol. 2009;169(4):505–14.
Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics. 2005;21(2):263–5.
Tregouet DA, Garelle V. A new JAVA interface implementation of THESIAS: testing haplotype effects in association studies. Bioinformatics. 2007;23(8):1038–9.
Penha-Goncalves C. Genetics of malaria inflammatory responses: a pathogenesis perspective. Front Immunol. 2019;10:1771.
Miossec P, Korn T, Kuchroo VK. Interleukin-17 and type 17 helper T cells. N Engl J Med. 2009;361(9):888–98.
Amezcua Vesely MC, Rodriguez C, Gruppi A, Acosta Rodriguez EV. Interleukin-17 mediated immunity during infections with Trypanosoma cruzi and other protozoans. Biochim Biophys Acta Mol Basis Dis. 2020;1866(5):165706.
Ishida H, Imai T, Suzue K, Hirai M, Taniguchi T, Yoshimura A, Iwakura Y, Okada H, Suzuki T, Shimokawa C, et al. IL-23 protection against Plasmodium berghei infection in mice is partially dependent on IL-17 from macrophages. Eur J Immunol. 2013;43(10):2696–706.
Fedorova L, Khrunin A, Khvorykh G, Lim J, Thornton N, Mulyar OA, Limborska S, Fedorov A. Analysis of common SNPs across continents reveals major genomic differences between human populations. Genes (Basel). 2022;13(8):1472.
Rahmah Z, Sasmito SD, Siswanto B, Sardjono TW, Fitri LE. Parasitemia induces high plasma levels of interleukin-17 (IL-17) and low levels of interleukin-10 (IL-10) and Transforming Growth Factor-ss (TGF-ss) in pregnant mice infected with malaria. Malays J Med Sci. 2015;22(3):25–32.
Herbert F, Tchitchek N, Bansal D, Jacques J, Pathak S, Becavin C, Fesel C, Dalko E, Cazenave PA, Preda C, et al. Evidence of IL-17, IP-10, and IL-10 involvement in multiple-organ dysfunction and IL-17 pathway in acute renal failure associated to Plasmodium falciparum malaria. J Transl Med. 2015;13:369.
Sahu U, Biswas D, Prajapati VK, Singh AK, Samant M, Khare P. Interleukin-17-A multifaceted cytokine in viral infections. J Cell Physiol. 2021;236(12):8000–19.
Scherer EF, Cantarini DG, Siqueira R, Ribeiro EB, Braga EM, Honorio-Franca AC, Franca EL. Cytokine modulation of human blood viscosity from vivax malaria patients. Acta Trop. 2016;158:139–47.
Tangteerawatana P, Pichyangkul S, Hayano M, Kalambaheti T, Looareesuwan S, Troye-Blomberg M, Khusmith S. Relative levels of IL4 and IFN-gamma in complicated malaria: association with IL4 polymorphism and peripheral parasitemia. Acta Trop. 2007;101(3):258–65.
Domingues W, Kanunfre KA, Rodrigues JC, Teixeira LE, Yamamoto L, Okay TS. Preliminary report on the putative association of Il10 -3575 T/a genetic polymorphism with malaria symptoms. Rev Inst Med Trop Sao Paulo. 2016;58:30.
Medina TS, Costa SP, Oliveira MD, Ventura AM, Souza JM, Gomes TF, Vallinoto AC, Povoa MM, Silva JS, Cunha MG. Increased interleukin-10 and interferon-gamma levels in Plasmodium vivax malaria suggest a reciprocal regulation which is not altered by IL-10 gene promoter polymorphism. Malar J. 2011;10:264.
Kaur R, Rawat AK, Kumar S, Aadil W, Akhtar T, Narang T, Chopra D. Association of genetic polymorphism of interleukin-17A & interleukin-17F with susceptibility of psoriasis. Indian J Med Res. 2018;148(4):422–6.
Zacarias JM, Sippert EA, Tsuneto PY, Visentainer JE, de Oliveira e Silva C, Sell AM. The influence of interleukin 17A and IL17F polymorphisms on chronic periodontitis disease in brazilian patients. Mediators Inflamm. 2015;2015:147056.
Marquet S, Conte I, Poudiougou B, Argiro L, Cabantous S, Dessein H, Burte F, Oumar AA, Brown BJ, Traore A, et al. The IL17F and IL17RA genetic variants increase risk of cerebral malaria in two African populations. Infect Immun. 2016;84(2):590–7.
Apinjoh TO, Anchang-Kimbi JK, Njua-Yafi C, Mugri RN, Ngwai AN, Rockett KA, Mbunwe E, Besingi RN, Clark TG, Kwiatkowski DP, et al. Association of cytokine and Toll-like receptor gene polymorphisms with severe malaria in three regions of Cameroon. PLoS One. 2013;8(11):e81071.
Israelsson E, Maiga B, Kearsley S, Dolo A, Homann MV, Doumbo OK, Troye-Blomberg M, Tornvall P, Berzins K. Cytokine gene haplotypes with a potential effect on susceptibility to malaria in sympatric ethnic groups in Mali. Infect Genet Evol. 2011;11(7):1608–15.
Okeyo WA, Munde EO, Okumu W, Raballah E, Anyona SB, Vulule JM, Ong’echa JM, Perkins DJ, Ouma C. Interleukin (IL)-13 promoter polymorphisms (-7402 T/G and -4729G/A) condition susceptibility to pediatric severe malarial anemia but not circulating IL-13 levels. BMC Immunol. 2013;14:15.
Phelan J, Gomez-Gonzalez PJ, Andreu N, Omae Y, Toyo-Oka L, Yanai H, Miyahara R, Nedsuwan S, de Sessions PF, Campino S, et al. Genome-wide host-pathogen analyses reveal genetic interaction points in tuberculosis disease. Nat Commun. 2023;14(1):549.
Mohanty S, Singh US, Mohanty S, Mohanty AK, Pande V, Das A. Evolutionary interplay of single nucleotide polymorphisms at the promoter region of TNF-alpha gene in different clinical outcomes of malaria in India. Infect Genet Evol. 2019;69:107–16.
Majewski J, Ott J. Distribution and characterization of regulatory elements in the human genome. Genome Res. 2002;12(12):1827–36.
Mucaki EJ, Shirley BC, Rogan PK. Expression changes confirm genomic variants predicted to result in allele-specific, alternative mRNA Splicing. Front Genet. 2020;11:109.
Dwyer K, Agarwal N, Pile L, Ansari A. Gene architecture facilitates intron-mediated enhancement of transcription. Front Mol Biosci. 2021;8:669004.
Robbez-Masson LJ, Bodor C, Jones JL, Hurst HC, Fitzgibbon J, Hart IR, Grose RP. Functional analysis of a breast cancer-associated FGFR2 single nucleotide polymorphism using zinc finger mediated genome editing. PLoS One. 2013;8(11):e78839.
Deng N, Zhou H, Fan H, Yuan Y. Single nucleotide polymorphisms and cancer susceptibility. Oncotarget. 2017;8(66):110635–49.
Faber K, Glatting KH, Mueller PJ, Risch A, Hotz-Wagenblatt A. Genome-wide prediction of splice-modifying SNPs in human genes using a new analysis pipeline called AASsites. BMC Bioinformatics. 2011;12 Suppl 4(Suppl 4):S2.
Lang X, Liu W, Hou Y, Zhao W, Yang X, Chen L, Yan Q, Cheng W. IL-17A polymorphism (rs2275913) and levels are associated with preeclampsia pathogenesis in Chinese patients. BMC Med Genomics. 2021;14(1):5.
Farag AGA, Labeeb AZ, Gerges ANA, Elshaib ME. Interleukin-17A in Egyptian leprosy patients: a clinical, genetic, and biochemical study. An Bras Dermatol. 2022;97(6):735–41.
Li N, Zhu Q, Li Z, Han Q, Zhang G, Chen J, Lv Y, Xing F, Chen Y, Zeng X, et al. IL17A gene polymorphisms, serum IL-17A and IgE levels, and hepatocellular carcinoma risk in patients with chronic hepatitis B virus infection. Mol Carcinog. 2014;53(6):447–57.
Lu F, Liu P, Zhang Q, Wang W, Guo W. Association between the polymorphism of IL-17A and IL-17F gene with knee osteoarthritis risk: a meta-analysis based on case-control studies. J Orthop Surg Res. 2019;14(1):445.
Acknowledgements
The authors are grateful to all the patients and the medical staff who have generously collaborated in the Malaria Genomic Project Clayton Dedonder 2014 (Institut Pasteur de Dakar). In addition, the authors thank the team of the Centre National de Recherche en Génétique humaine (CNRGH) for assistance in performing the genotyping. The French Ministry of Education and Research supported the CNRGH-CEA.
Funding
This research received no external funding.
Author information
Authors and Affiliations
Contributions
FT conceived the study and the methodology and drafted the manuscript. GD conceived the study and the methodology. JFZ and CC conducted the data analysis, revised the manuscript and approved the final version. CD conducted the methodology, performed Sanger Sequencing and molecular biology experiments and approved the final version. BM conducted the malaria cohort recruitment, revised the manuscript from the hospital center, and approved the final version. AAMD, AT, MD, CMN, YD contributed to the correction and revision of the manuscript and approved the final version. JFD & AD coordinated this study, revised the manuscript, and approved the final version. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
“The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee of UCAD’s Committee on Research and Ethics (CER) considers that the research proposed respects the appropriate ethical standard and, as a result, approves its execution under “Protocole 0344/2018/CER-UCAD”. Written informed consent was obtained from adult participants and parents or legal representatives of children and also was obtained from the patient(s) to publish this paper.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Thiam, F., Diop, G., Coulonges, C. et al. An elevated level of interleukin-17A in a Senegalese malaria cohort is associated with rs8193038 IL-17A genetic variant. BMC Infect Dis 24, 275 (2024). https://doi.org/10.1186/s12879-024-09149-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12879-024-09149-8